
Collateral Effects of Insecticide-Treated Nets on Human and Environmental Safety in an Epidemiological Model for Malaria with Human Risk Perception

Abstract
:1. Introduction
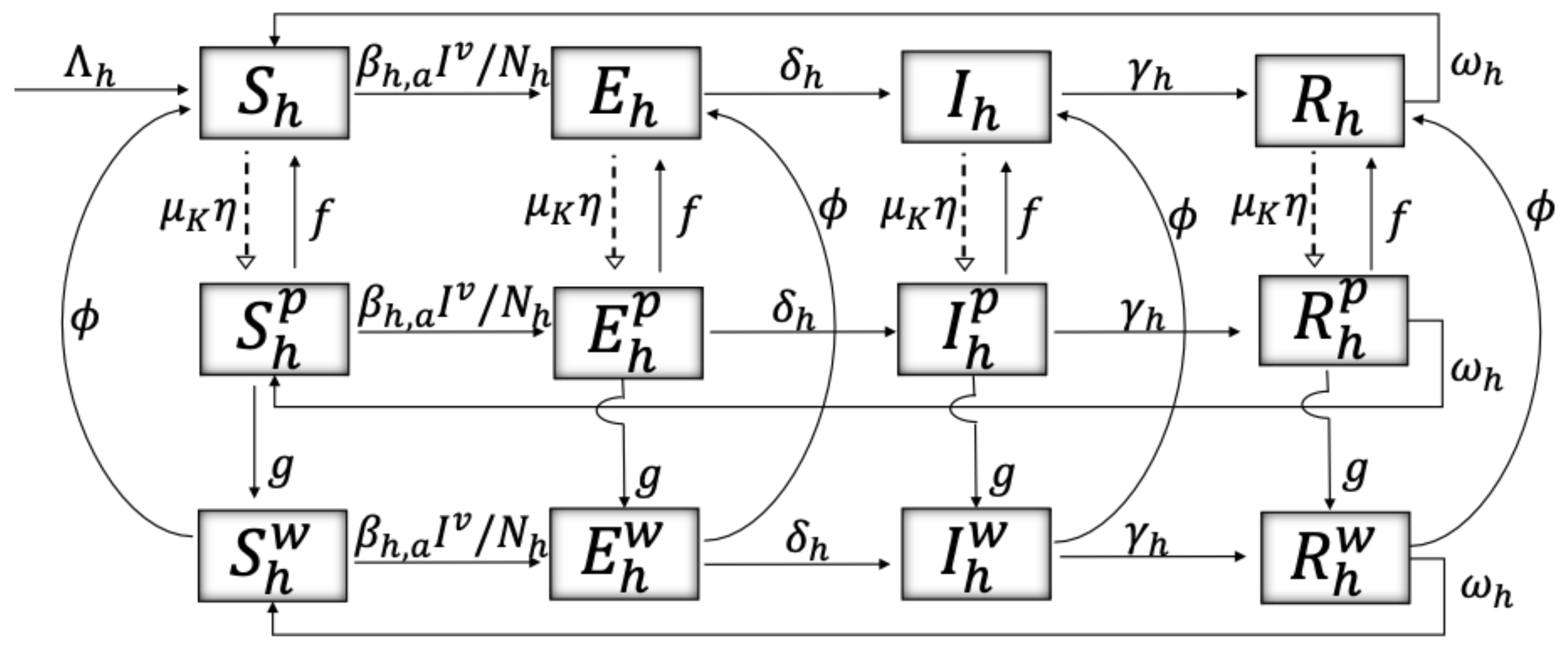
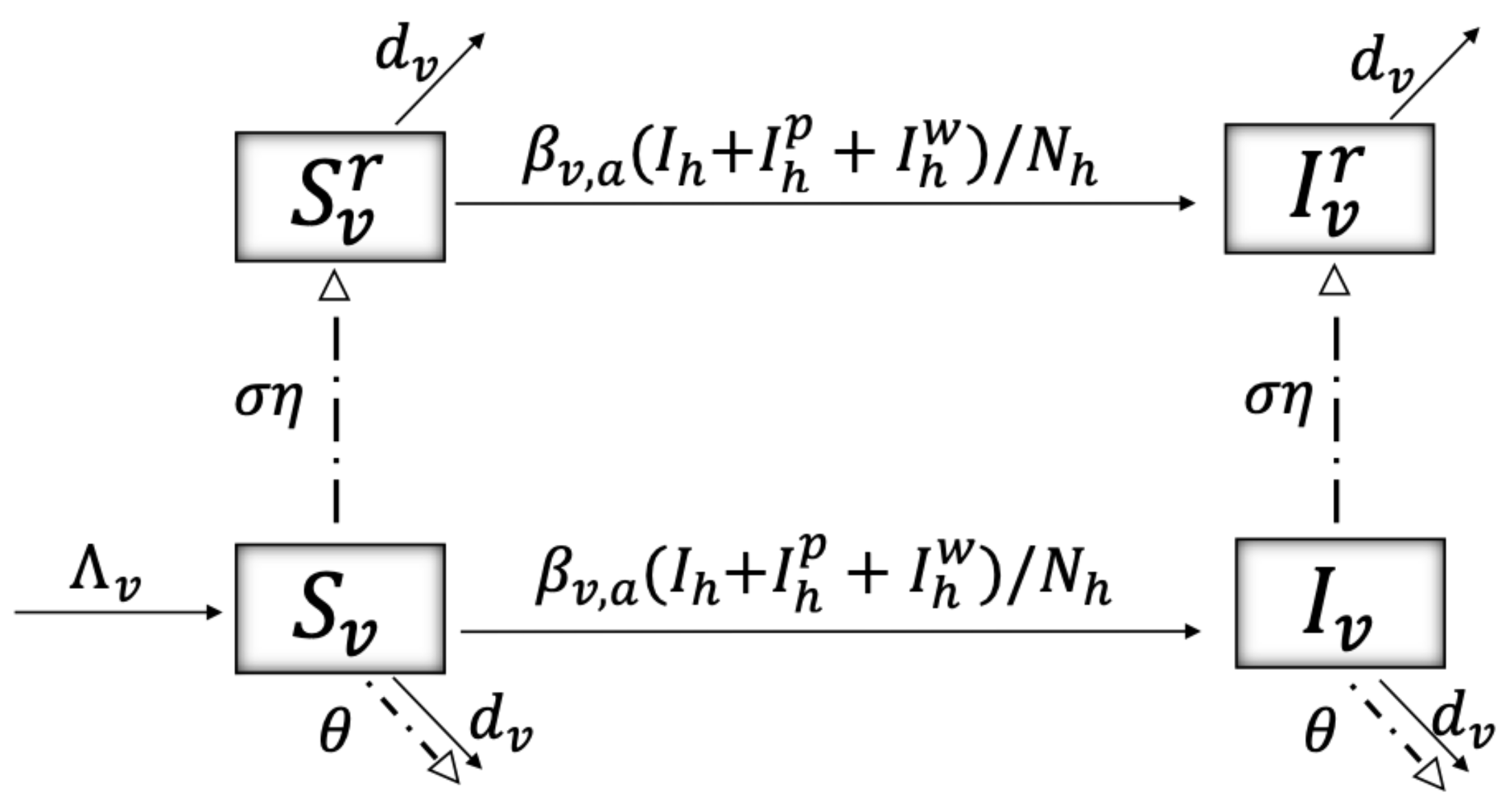
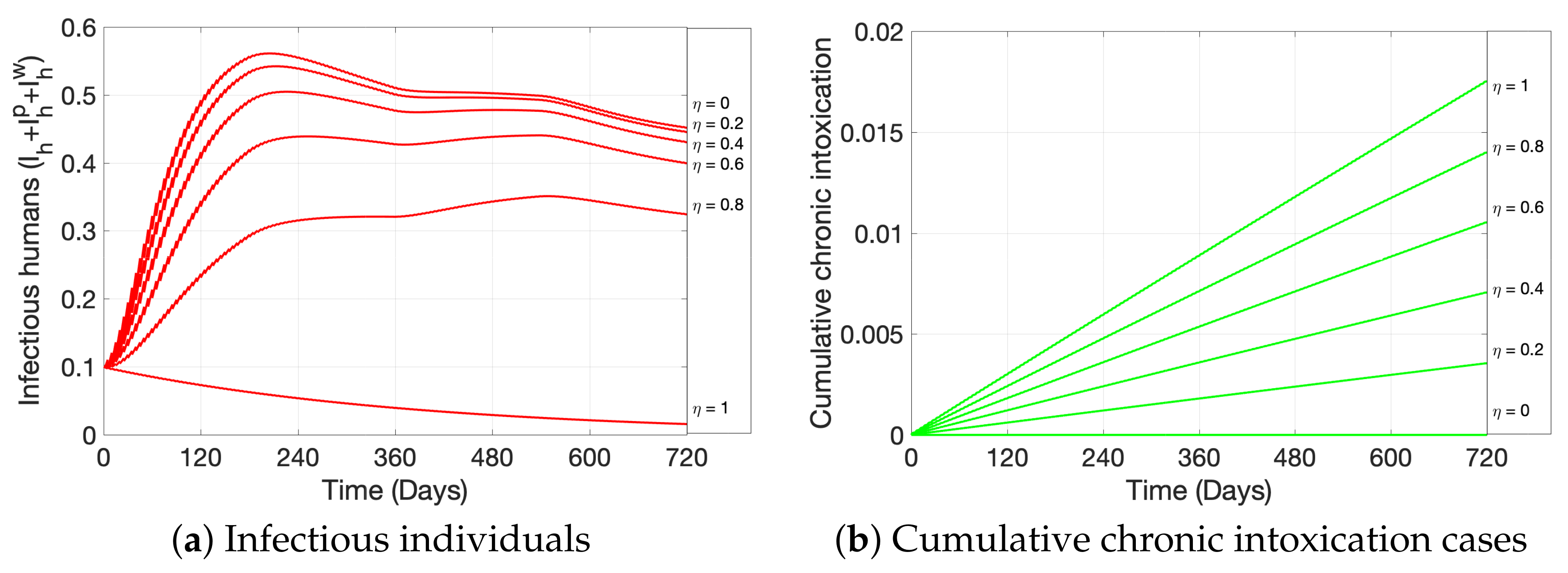
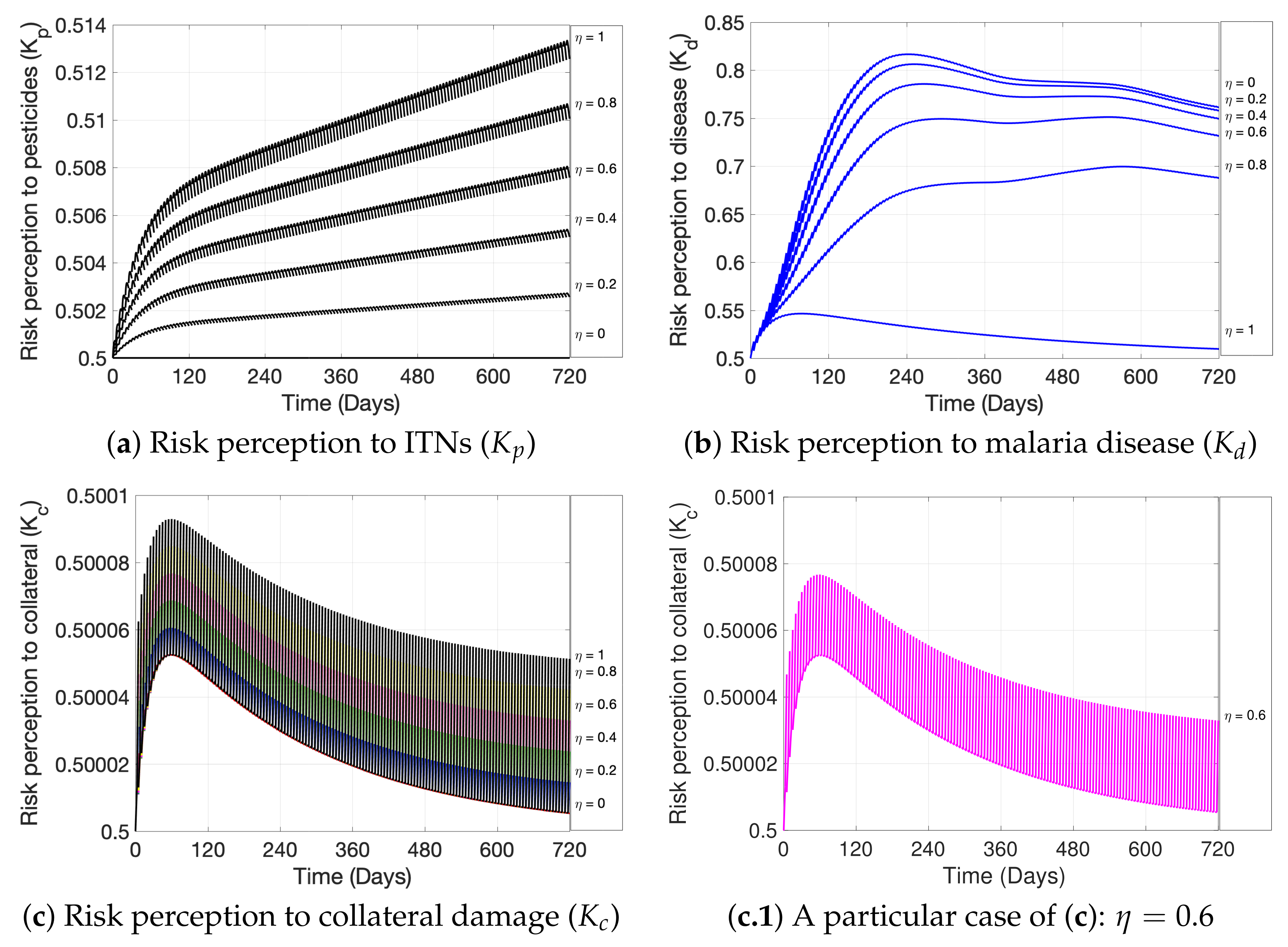
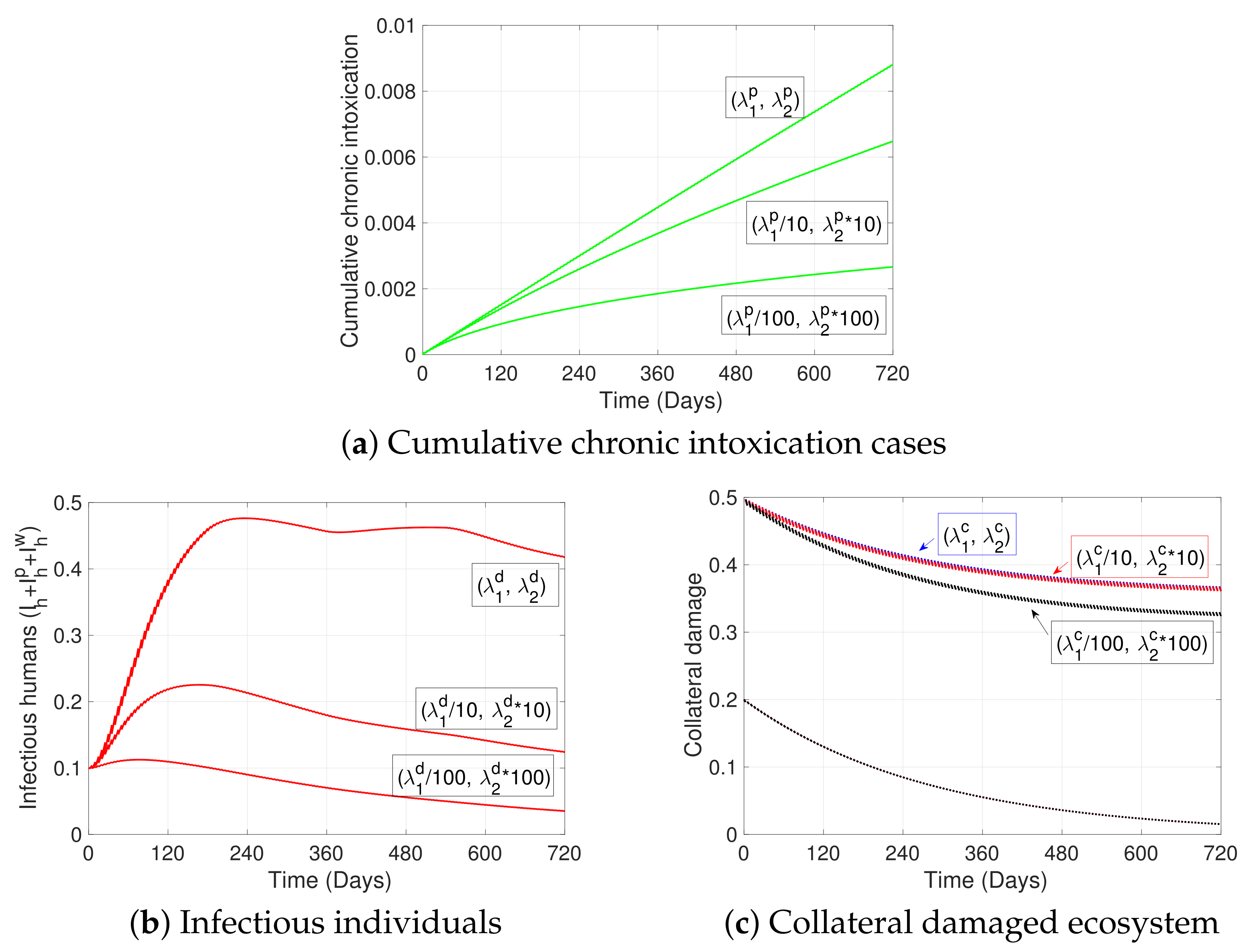
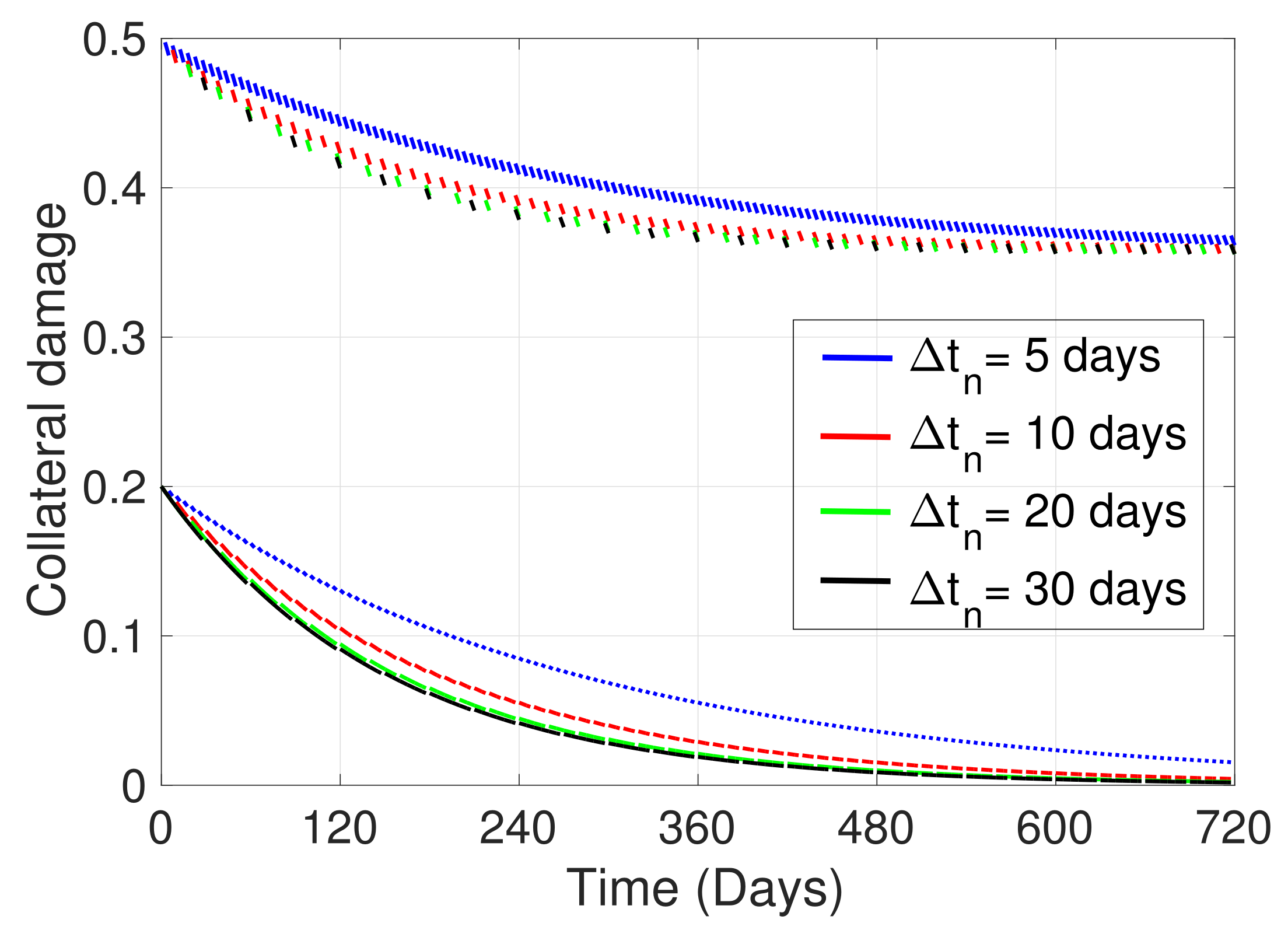
2. Materials and Methods
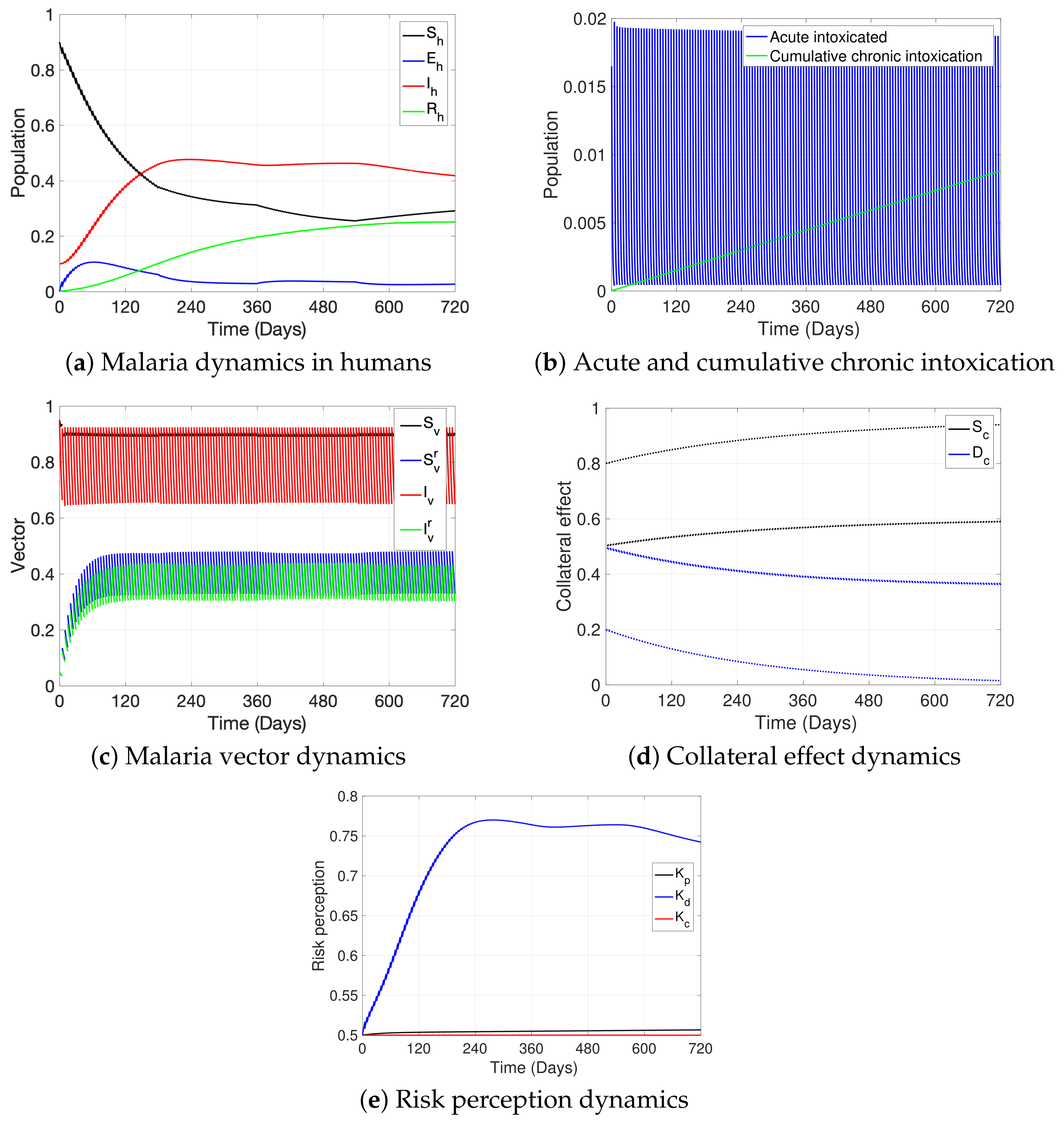
3. Simulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ITN | Insecticide-treated net |
| WHO | World Health Organization |
References
- World Health Organization (WHO). Malaria. Available online: https://www.who.int/news-room/fact-sheets/detail/malaria (accessed on 3 October 2022).
- Centers for Disease Control and Prevention (CDC). About Malaria. Available online: https://www.cdc.gov/malaria/about/ (accessed on 23 August 2022).
- Almaw, A.; Yimer, M.; Alemu, M.; Tegegne, B. Prevalence of malaria and associated factors among symptomatic pregnant women attending antenatal care at three health centers in north-west Ethiopia. PLoS ONE 2022, 17, e0266477. [Google Scholar] [CrossRef]
- Uneke, C.J.; Ogbu, O.; Nwojiji, V. Potential risk of induced malaria by blood transfusion in South-eastern Nigeria. McGill J. Med. MJM 2006, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.; Taylor, G.; Saldaña-Núñez, V.; Córdova-Lepe, F.; Escalera-Antezana, J.P.; Alvarado-Arnez, L.E.; Rodríguez-Morales, A.J. Risk of Dengue Incidence in Children and Adolescents in Zulia, Venezuela, using a Negative Binomial Generalized Linear Mixed Model. Rev. Panam. Enferm. Infecc. 2019, 2, 39–45. [Google Scholar]
- Guyatt, H.L.; Snow, R.W. Impact of malaria during pregnancy on low birth weight in sub-Saharan Africa. Clin. Microbiol. Rev. 2004, 17, 760–769. [Google Scholar] [CrossRef] [Green Version]
- Roll Back Malaria. Available online: https://endmalaria.org/about-malaria/key-facts (accessed on 30 July 2022).
- World Health Organization (WHO). World Malaria Report 2021. Available online: https://www.who.int/publications/i/item/9789240040496 (accessed on 3 October 2022).
- Pryce, J.; Medley, N.; Choi, L. Indoor residual spraying for preventing malaria in communities using insecticide-treated nets. Cochrane Database Syst. Rev. 2022, 5, CD012688. [Google Scholar]
- Sherrard-Smith, E.; Winskill, P.; Hamlet, A.; Ngufor, C.; N’Guessan, R.; Guelbeogo, M.W.; Sanou, A.; Nash, R.K.; Hill, A.; Russell, E.L.; et al. Optimising the deployment of vector control tools against malaria: A data-informed modelling study. Lancet Planet. Health 2022, 6, e100–e109. [Google Scholar] [CrossRef] [PubMed]
- Pryce, J.; Richardson, M.; Lengeler, C. Insecticide-treated nets for preventing malaria. Cochrane Database Syst. Rev. 2018, 11, CD000363. [Google Scholar] [CrossRef]
- Wetzler, E.A.; Park, C.; Arroz, J.A.; Chande, M.; Mussambala, F.; Candrinho, B. Impact of mass distribution of insecticide-treated nets in Mozambique, 2012 to 2025: Estimates of child lives saved using the Lives Saved Tool. PLoS Glob. Public Health 2022, 2, e0000248. [Google Scholar] [CrossRef]
- Millat-Martínez, P.; Gabong, R.; Balanza, N.; Luana, S.; Sanz, S.; Raulo, S.; Elizah, A.; Wali, C.; Paivu, B.; Dalmas, J.; et al. Coverage, determinants of use and repurposing of long-lasting insecticidal nets two years after a mass distribution in Lihir Islands, Papua New Guinea: A cross-sectional study. Malar. J. 2021, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Larsen, D.A.; Makaure, J.; Ryan, S.J.; Stewart, D.; Traub, A.; Welsh, R.; Love, D.H.; Bisesi, J.H., Jr. Implications of insecticide-treated mosquito net fishing in lower income countries. Environ. Health Perspect. 2021, 129, 015001. [Google Scholar] [CrossRef]
- McLean, K.A.; Byanaku, A.; Kubikonse, A.; Tshowe, V.; Katensi, S.; Lehman, A.G. Fishing with bed nets on Lake Tanganyika: A randomized survey. Malar. J. 2014, 13, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, R.; Gurung, R.; Rowcliffe, M.; Hill, N.; Milner-Gulland, E. The use of mosquito nets in fisheries: A global perspective. PLoS ONE 2018, 13, e0191519. [Google Scholar] [CrossRef]
- Berthe, S.; Harvey, S.A.; Lynch, M.; Koenker, H.; Jumbe, V.; Kaunda-Khangamwa, B.; Mathanga, D.P. Poverty and food security: Drivers of insecticide-treated mosquito net misuse in Malawi. Malar. J. 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Jones, B.L.; Unsworth, R.K. The perverse fisheries consequences of mosquito net malaria prophylaxis in East Africa. Ambio 2020, 49, 1257–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Guidelines for Malaria. 2022. Available online: https://apps.who.int/iris/bitstream/handle/10665/354781/WHO-UCN-GMP-2022.01-Rev.2-eng.pdf (accessed on 4 October 2022).
- RBM Partnership to End Malaria, Consensus Statement on Repurposing ITNs: Applications for BCC Messaging and Actions at the Country Level. 2018. Available online: https://pdf.usaid.gov/pdf_docs/PA00TW8X.pdf (accessed on 2 October 2022).
- USAID Programmatic Environmental Assessment: Integrated Vector Management Programs for Malaria Vector Control (version 2017). Available online: https://d1u4sg1s9ptc4z.cloudfront.net/uploads/2021/03/integrated-vector-management-programs-for-malaria-vector-control-programmatic-environmental-assessment-2017-2.pdf (accessed on 4 October 2022).
- Hu, R.; Huang, X.; Huang, J.; Li, Y.; Zhang, C.; Yin, Y.; Chen, Z.; Jin, Y.; Cai, J.; Cui, F. Long- and Short-Term Health Effects of Pesticide Exposure: A Cohort Study from China. PLoS ONE 2015, 10, 128766. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.; Kim, S.K.; Kim, C.S.; Kim, T.H.; Min, S.H.; Oh, S.S.; Koh, S.B. Correction: Pesticide exposure and cognitive decline in a rural South Korean population. PLoS ONE 2019, 14, 216310. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Schelar, E. Pesticide Exposure and Child Neurodevelopment: Summary and Implications. Workplace Health Saf. 2012, 60, 235–242. [Google Scholar] [CrossRef]
- Muñoz-Quezada, M.T.; Lucero, B.A.; Gutiérrez-Jara, J.P.; Buralli, R.J.; Zúñiga-Venegas, L.; Muñoz, M.P.; Ponce, K.V.; Iglesias, V. Longitudinal exposure to pyrethroids (3-PBA and trans-DCCA) and 2,4-D herbicide in rural schoolchildren of Maule region, Chile. Sci. Total Environ. 2020, 749, 141512. [Google Scholar] [CrossRef]
- Zúñiga-Venegas, L.A.; Hyland, C.; Muñoz-Quezada, M.T.; Quirós-Alcalá, L.; Butinof, M.; Buralli, R.; Cardenas, A.; Fernandez, R.A.; Foerster, C.; Gouveia, N.; et al. Health Effects of Pesticide Exposure in Latin American and the Caribbean Populations: A Scoping Review. Environ. Health Perspect. 2022, 130, 096002. [Google Scholar] [CrossRef]
- Anyanwu, E.C.; Ehiri, J.E.; Kanu, I.; Merrick, J. Health effects of long-term exposure to insecticide-treated mosquito nets in the control of malaria in endemic regions, revised. Sci. World J. 2006, 6, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Kolaczinski, J.; Curtis, C. Chronic illness as a result of low-level exposure to synthetic pyrethroid insecticides: A review of the debate. Food Chem. Toxicol. 2004, 42, 697–706. [Google Scholar] [CrossRef]
- Bomann, W. How safe are pyrethroid-treated mosquito nets? An evaluation based on the example of Solfac EW 050. Public Health 1995, 12, 30–35. [Google Scholar]
- Lu, G.; Traoré, C.; Meissner, P.; Kouyaté, B.; Kynast-Wolf, G.; Beiersmann, C.; Coulibaly, B.; Becher, H.; Müller, O. Safety of insecticide-treated mosquito nets for infants and their mothers: Randomized controlled community trial in Burkina Faso. Malar. J. 2015, 14, 1–7. [Google Scholar] [CrossRef]
- Brauer, F.; Castillo-Chavez, C.; Mubayi, A.; Towers, S. Some models for epidemics of vector-transmitted diseases. Infect. Dis. Model. 2016, 1, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Simoy, M.I.; Aparicio, J.P. Ross-Macdonald models: Which one should we use? Acta Trop. 2020, 207, 105452. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Jara, J.P.; Salazar-Viedma, M.; González, C.R.; Cancino-Faure, B. The emergence of Dirofilaria repens in a non-endemic area influenced by climate change: Dynamics of transmission using a mathematical model. Acta Trop. 2022, 226, 106230. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. Some a priori pathometric equations. Br. Med. J. 1915, 1, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngonghala, C.N. Assessing the impact of insecticide-treated nets in the face of insecticide resistance on malaria control. J. Theor. Biol. 2022, 555, 111281. [Google Scholar] [CrossRef]
- Ndamuzi, E.; Gahungu, P. Mathematical Modeling of Malaria Transmission Dynamics: Case of Burundi. J. Appl. Math. Phys. 2021, 9, 2447–2460. [Google Scholar] [CrossRef]
- Witbooi, P.; Abiodun, G.; Nsuami, M. A model of malaria population dynamics with migrants. Math. Biosci. Eng. 2021, 18, 7301–7317. [Google Scholar] [CrossRef] [PubMed]
- Vogt-Geisse, K.; Ngonghala, C.N.; Feng, Z. The impact of vaccination on malaria prevalence: A vaccine-age-structured modeling approach. J. Biol. Syst. 2020, 28, 475–513. [Google Scholar] [CrossRef]
- Vogt-Geisse, K.; Lorenzo, C.; Feng, Z. Impact of age-dependent relapse and immunity on malaria dynamics. J. Biol. Syst. 2013, 21, 1340001. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Sarkar, R.R.; Sinha, S. Mathematical models of malaria-a review. Malar. J. 2011, 10, 1–19. [Google Scholar] [CrossRef]
- Lindsay, S.W.; Thomas, M.B.; Kleinschmidt, I. Threats to the effectiveness of insecticide-treated bednets for malaria control: Thinking beyond insecticide resistance. Lancet Glob. Health 2021, 9, e1325–e1331. [Google Scholar] [CrossRef] [PubMed]
- White, L.J.; Maude, R.J.; Pongtavornpinyo, W.; Saralamba, S.; Aguas, R.; Van Effelterre, T.; Day, N.P.; White, N.J. The role of simple mathematical models in malaria elimination strategy design. Malar. J. 2009, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitnis, N.; Schapira, A.; Smith, T.; Steketee, R. Comparing the effectiveness of malaria vector-control interventions through a mathematical model. Am. J. Trop. Med. Hyg. 2010, 83, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agusto, F.B.; Del Valle, S.Y.; Blayneh, K.W.; Ngonghala, C.N.; Goncalves, M.J.; Li, N.; Zhao, R.; Gong, H. The impact of bed-net use on malaria prevalence. J. Theor. Biol. 2013, 320, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Chinebu, T.I.; Ezennorom, E.O.; Okwor, J.U. Simulation of a Mathematical Model of Malaria Transmission Dynamics in the Presence of Mosquito Net, Fumigation And Treatment. Int. J. Trend Sci. Res. Dev. 2018, 2, 339–360. [Google Scholar] [CrossRef] [Green Version]
- Aldila, D. Dynamical Analysis on a Malaria Model with Relapse Preventive Treatment and Saturated Fumigation. Comput. Math. Methods Med. 2022, 2022, 1135452. [Google Scholar] [CrossRef]
- Córdova-Lepe, F.; Pinto, M.; González-Olivares, E. A new class of differential equations with impulses at instants dependent on preceding pulses. Applications to management of renewable resources. Nonlinear Anal. Real World Appl. 2012, 13, 2313–2322. [Google Scholar] [CrossRef]
- Liu, H.; Yu, J.; Zhu, G. Analysis of a vector-host malaria model with impulsive effect and infection-age. Adv. Complex Syst. 2006, 9, 237–248. [Google Scholar] [CrossRef]
- Huang, M.; Song, X.; Li, J. Modelling and analysis of impulsive releases of sterile mosquitoes. J. Biol. Dyn. 2017, 11, 147–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Jara, J.P.; Córdova-Lepe, F.; Muñoz-Quezada, M.T.; Chowell, G. Pesticide application, educational treatment and infectious respiratory diseases: A mechanistic model with two impulsive controls. PLoS ONE 2020, 15, 243048. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.; Cordova-Lepe, F.; Gutierrez-Jara, J.P.; Vogt Geisse, K. An SIR type epidemiological model that integrates social distancing as a dynamic law based on point prevalence and socio-behavioral factors. Sci. Rep. 2021, 11, 10170. [Google Scholar] [CrossRef]
- Gutiérrez-Jara, J.P.; Vogt Geisse, K.; Cabrera, M.; Cordova-Lepe, F.; Muñoz-Quezada, M.T. Effects of human mobility and behavior on disease transmission in a COVID-19 mathematical model. Sci. Rep. 2022, 12, 10840. [Google Scholar] [CrossRef]
- Gutiérrez-Jara, J.P.; Saracini, C. Risk Perception Influence on Vaccination Program on COVID-19 in Chile: A Mathematical Model. Int. J. Environ. Res. Public Health 2022, 19, 2022. [Google Scholar] [CrossRef]
- Chitnis, N.; Hyman, M.J.; Cushing, J.M. Determining Important Parameters in the Spread of Malaria Through the Sensitivity Analysis of a Mathematical Model. Bull. Math. Biol. 2008, 70, 1272–1296. [Google Scholar] [CrossRef]
- Nah, K.; Kim, Y.; Lee, J.M. The dilution effect of the domestic animal population on the transmission of P. vivax malaria. J. Theor. Biol. 2010, 266, 299–306. [Google Scholar] [CrossRef]
- de Alencar, F.E.; Malafronte, R.D.S.; Cerutti Junior, C.; Natal Fernandes, L.; Buery, J.C.; Fux, B.; Rezende, H.R.; Duarte, A.M.R.D.C.; Medeiros-Sousa, A.R.; Miranda, A.E. Assessment of asymptomatic Plasmodium spp. infection by detection of parasite DNA in residents of an extra-Amazonian. Malar. J. 2021, 17, 1475–2875. [Google Scholar] [CrossRef] [Green Version]
- Medeiros-Sousa, A.R.; Laporta, G.Z.; Coutinho, R.M.; Mucci, L.F.; Marrelli, M.T. A mathematical model for zoonotic transmission of malaria in the Atlantic Forest: Exploring the effects of variations in vector abundance and acrodendrophily. PLoS Negl. Trop. Dis. 2021, 15, 1–21. [Google Scholar] [CrossRef]
- Kim, S.; Byun, J.H.; Park, A.; Jung, I.H. A mathematical model for assessing the effectiveness of controlling relapse in Plasmodium vivax malaria endemic in the Republic of Korea. PLoS ONE 2020, 15, 227919. [Google Scholar] [CrossRef] [PubMed]
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxical Profile for Pyrethrins and Pyrethroids. Available online: https://www.atsdr.cdc.gov/toxprofiledocs/index.html?id=787&tid=153 (accessed on 5 August 2022).
- Rodríguez, J.L.; Ares, I.; Martínez, M.; Martínez-Larrañaga, M.R.; Anadón, A.; Martínez, M.A. Bioavailability and nervous tissue distribution of pyrethroid insecticide cyfluthrin in rats. Food Chem. Toxicol. 2018, 118, 220–226. [Google Scholar] [CrossRef] [PubMed]
- World Bank Data, Life Expectancy at Birth, Total (Years)—Sub-Saharan Africa. Available online: https://data.worldbank.org/indicator/SP.DYN.LE00.IN?locations=ZG (accessed on 15 August 2022).
- The World Factbook, Central Intelligence Agency. Life Expectancy at Birth. Available online: https://www.cia.gov/the-world-factbook/field/life-expectancy-at-birth/country-comparison (accessed on 14 August 2022).
- Insecticide-Treated Mosquito Net Interventions: A Manual for National Control Programme Managers. Edited by Roll Back Malaria. Available online: https://apps.who.int/iris/handle/10665/42685 (accessed on 15 August 2022).
- Laporta, G.Z.; Prado, P.I.K.L.d.; Kraenkel, R.A.; Coutinho, R.M.; Sallum, M.A.M. Biodiversity Can Help Prevent Malaria Outbreaks in Tropical Forests. PLoS Negl. Trop. Dis. 2013, 7, e2139. [Google Scholar] [CrossRef]
- Kouassi, B.L.; Edi, C.; Tia, E.; Konan, L.Y.; Akré, M.A.; Koffi, A.A.; Ouattara, A.F.; Tanoh, A.M.; Zinzindohoue, P.; Kouadio, B.; et al. Susceptibility of Anopheles gambiae from Côte d’Ivoire to insecticides used on insecticide-treated nets: Evaluating the additional entomological impact of piperonyl butoxide and chlorfenapyr. Malar. J. 2020, 19, 454. [Google Scholar] [CrossRef] [PubMed]
- Mwandya, A.W.; Gullström, M.; Öhman, M.C.; Andersson, M.H.; Mgaya, Y.D. Fish assemblages in Tanzanian mangrove creek systems influenced by solar salt farm constructions. Estuar. Coast. Shelf Sci. 2009, 82, 193–200. [Google Scholar] [CrossRef]
- El Ganainy, A.; Khouraiba, H.; Aly, M.; Osman, M.A. Fishery assessment of the common silver biddy gerres oyena from the gulf of Suez, Red Sea, Egypt. Egypt. J. Aquat. Biol. Fish. 2020, 24, 437–447. [Google Scholar] [CrossRef]
- Yongo, E.; Agembe, S.; Outa, N.; Owili, M. Growth, mortality and recruitment of Nile perch (Lates niloticus) in Lake Victoria, Kenya. Lakes Reserv. Res. Manag. 2018, 23, 17–23. [Google Scholar] [CrossRef]
- Owusu, V.; Andriesse, E. From open access regime to closed fishing season: Lessons from small-scale coastal fisheries in the Western Region of Ghana. Mar. Policy 2020, 121, 104162. [Google Scholar] [CrossRef]
- Cabrera, M.; Taylor, G. Modelling spatio-temporal data of dengue fever using generalized additive mixed models. Spat. Spatio-Temporal Epidemiol. 2019, 28, 1–13. [Google Scholar] [CrossRef]
- Córdova-Lepe, F.; Vogt-Geisse, K. Adding a reaction-restoration type transmission rate dynamic-law to the basic SEIR COVID-19 model. PLoS ONE 2022, 17, e0269843. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
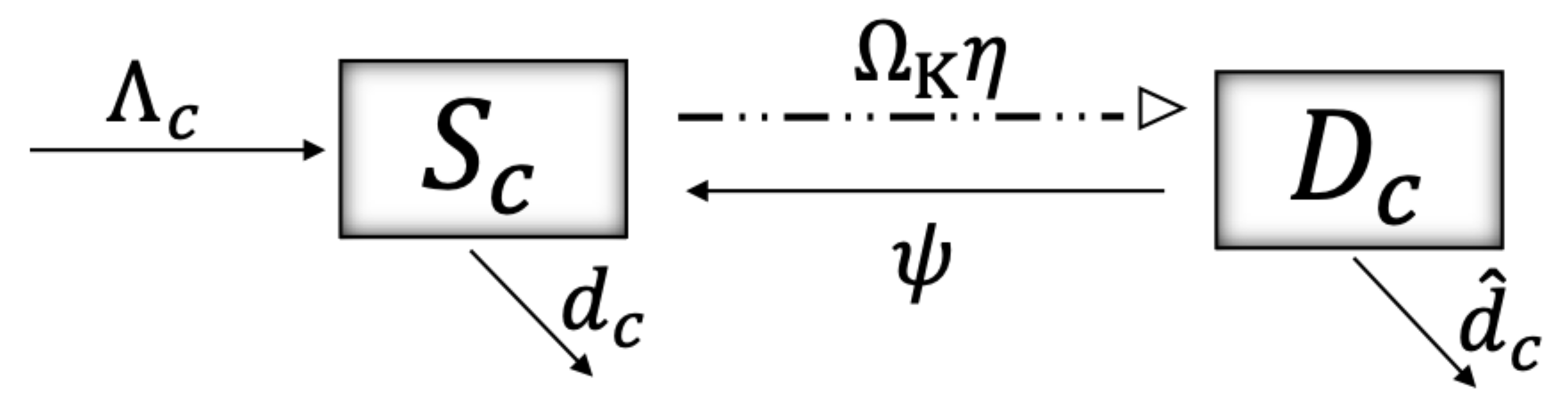{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Definition | Units * |
|---|---|---|
| Humans | ||
| Malaria-susceptible individuals w/o, w/ acute, w/ chronic intoxication | H | |
| Malaria-exposed individuals w/o, w/ acute, w/ chronic intoxication | H | |
| Malaria-infected individuals w/o, w/ acute, w/ chronic intoxication | H | |
| Malaria-recovered individuals w/o, w/ acute, w/ chronic intoxication | H | |
| Risk perception to ITNs toxicity | R | |
| Risk perception to malaria disease | R | |
| Risk perception to collateral ecosystem damage from misuse of ITNs | R | |
| Vector | ||
| Malaria susceptible mosquito w/o, w/ insecticide resistance | M | |
| Malaria infected mosquito w/o, w/ insecticide resistance | M | |
| Collateral effect—aquatic ecosystem | ||
| Ecosystem susceptible to damage | C | |
| Damaged ecosystem | C | |
| Parameter | Definition | Baseline | Units * | Reference |
|---|---|---|---|---|
| Humans | ||||
| Malaria transmission rate to human | [0.015, 0.22] | D | [54,55,56,57,58] | |
| Average incubation time | [12, 30] | D | [55,58] | |
| Mean infectious period | [180, 720] | D | [54,55,56,57,58] | |
| Loss of immunity rate | D | [54] | ||
| Intoxication proportion | [0, 0.3] | U | [26,30] | |
| Mean detoxification period | [1/6, 2] | D | [59,60] | |
| g | Health effects after prolonged exposure | 0.00289 | D | [25,26] |
| Recovery rate from chronic toxicity | [0, 0.5] | D | Author chosen | |
| Recruitment rate | HD | Author chosen | ||
| Natural mortality rate | D | [61,62] | ||
| Disease-induced mortality rate | D | [54] | ||
| Rate of resistance to change risk perception | [0, 1] | D | [25,51,52,53] | |
| Per capita reaction to change risk perception | [0, 1] | RD, R | [25,51,52,53] | |
| Mosquito net coverage | [0, 1] | U | [7] | |
| Vector | ||||
| Malaria transmission rate to mosquito | [0.015, 0.24] | D | [54,55,56,57,58] | |
| Proportion of mosquitoes becoming resistant | [0, 1] | U | [1,7,63] | |
| Recruitment rate | MD | Author chosen | ||
| Natural mortality rate | [1/30, 1] | D | [57,64] | |
| Increase in the mortality rate | [4.4, 48.9]% | U | [65] | |
| Collateral effect—aquatic ecosystem | ||||
| Percentage of species susceptible to ITN fishing | U | [66] | ||
| Recruitment rate | CD | Author chosen | ||
| Natural mortality rate | D | [67,68] | ||
| Damage-induced mortality rate | D | [67,68] | ||
| Loss of damage rate | D | [67,68] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Jara, J.P.; Vogt-Geisse, K.; Cabrera, M. Collateral Effects of Insecticide-Treated Nets on Human and Environmental Safety in an Epidemiological Model for Malaria with Human Risk Perception. Int. J. Environ. Res. Public Health 2022, 19, 16327. https://doi.org/10.3390/ijerph192316327
Gutiérrez-Jara JP, Vogt-Geisse K, Cabrera M. Collateral Effects of Insecticide-Treated Nets on Human and Environmental Safety in an Epidemiological Model for Malaria with Human Risk Perception. International Journal of Environmental Research and Public Health. 2022; 19(23):16327. https://doi.org/10.3390/ijerph192316327
Chicago/Turabian StyleGutiérrez-Jara, Juan Pablo, Katia Vogt-Geisse, and Maritza Cabrera. 2022. "Collateral Effects of Insecticide-Treated Nets on Human and Environmental Safety in an Epidemiological Model for Malaria with Human Risk Perception" International Journal of Environmental Research and Public Health 19, no. 23: 16327. https://doi.org/10.3390/ijerph192316327