





Abstract
Astronomers consider light pollution to be a growing problem, however few studies have addressed potential effects of light pollution on wildlife. Sunlight is believed to initiate song in many bird species. If light initiates song, then light pollution may be influencing avian song behavior at a population level. This hypothesis predicts that birds breeding in areas with large amounts of artificial light will begin singing earlier in the day than birds in areas with little artificial light. Birds in highly illuminated areas might begin singing earlier than did birds in those same areas in previous years when artificial light levels were known to be, or were presumably, lower. Also, birds should begin singing earlier within a site on brightly lit nights. In 2002 and 2003 I documented initiation of morning song by breeding American Robins (Turdus migratorius) in areas with differing intensity of artificial nocturnal light. I compared my observations among sites and against historical studies. Robin populations in areas with large amounts of artificial light frequently began their morning chorus during true night. Chorus initiation time, relative to civil twilight, was positively correlated with amount of artificial light present during true night. Robin choruses in areas with little, or presumably little, artificial light have almost never begun during true night, instead appearing to track the onset of civil twilight. Proliferation of artificial nocturnal light may be strongly affecting singing behavior of American Robins at a population level.
Resumen
Efectos Aparentes de la Polución Luminica Sobre el Comportamiento del Canto en Turdus migratorius
Los astrónomos consideran que la polución luminica es un problema creciente, aunque pocos estudios han evaluado sus efectos potenciales en la fauna silvestre. Se cree que la luz del sol estimula el inicio del canto en muchas especies de aves. Si la luz inicia el canto, entonces la polución luminica puede estar influenciando el comportamiento del canto a nivel poblacional. Esta hipótesis predice que las aves que crian en áreas con altos niveles de luz artificial comenzarán a cantar más temprano en el dia que aquellas en áreas con poca luz artificial. Las aves de áreas altamente iluminadas podrian comenzar a cantar más temprano que las aves en las mismas áreas en los años previos cuando los niveles de luz artificial eran, o se presumia que eran, menores. De igual manera, las aves deberian comenzar a cantar más temprano en los sitios con noches luminosas. En el 2002 y el 2003, documenté el inicio de los cantos matutinos de Turdus migratorius en áreas con diferentes intensidades de luz artificial nocturna. Comparé mis observaciones entre sitios y con estudios previos. Las poblaciones de T. migratorius en las áreas con niveles elevados de luz artificial frecuentemente comenzaron sus coros matutinos durante la noche real. El tiempo de inicio del coro, en relación con el anochecer urbano, se correlacionó positivamente con la cantidad de luz artificial presente durante la noche. Los coros de T. migratorius en las áreas con poca, o presumiblemente poca, luz artificial casi nunca han comenzado durante la noche real y en su lugar aparentan seguir el inicio del anochecer urbano. La proliferación de la luz artificial nocturna puede estar afectando severamente el comportamiento del canto de T. migratorius a nivel poblacional.
Introduction
Many bird species begin singing near dawn. Quantitative study of dawn singing began in the late 19th and early 20th centuries, providing evidence that sunlight initiates bird song (Wright 1912, Allen 1913, Allard 1930). Bright moonlight may cause birds to begin morning song earlier than normal (Leopold and Eynon 1961), while cloud cover may delay the start of song (Allard 1930, Emlen 1937). However, effect of light level on bird song initiation has never been directly evaluated. If light initiates avian song then song initiation times, relative to civil twilight, should correlate positively with morning light levels within and among sites.
Light pollution is frequently identified by astronomers as a growing problem (Berry 1976, Crelin 2002, Tyson 2002); however, possible effects of light pollution on wildlife populations have received little study (Bower 2000, Harder 2002, Le Corre et al. 2002). If light initiates bird song and light pollution affects singing behavior at a population level, then birds breeding in areas with large amounts of artificial light should begin singing earlier in the day than birds in areas with little artificial light. Birds in highly illuminated areas might also begin singing earlier than did birds in those same areas in previous years when artificial light levels were known to be, or were presumably, lower. Also, daily song initiation times within a site should correlate positively with light levels recorded during true night. I examined these predictions by recording when American Robins (Turdus migratorius) began their morning chorus at several sites differing in level of artificial light, and by comparing those times with previous studies conducted in the early–mid 20th century (Allard 1930, Leopold and Eynon 1961). I chose to study the American Robin because it is common in urban and rural habitats (Sallabanks and James 1999) and exhibits a strong morning chorus, but does not normally sing during true night (Allard 1930, Leopold and Eynon 1961, Fisler 1962).
Methods
Pennsylvania Study Areas
In 2002 three field sites were studied in Schuylkill County, Pennsylvania. Sites were selected for obvious differences in levels of artificial light, and for geographic proximity. All three sites were in the valley-and-ridge physiographic province. Distance and mountainous terrain obscured artificial light among sites.
Schuylkill Haven, a small city (pop. 5548, 40°38′N, 76°11′W), had a large amount of artificial light. The study area was dominated by row houses and a foundry. Over 100 outdoor lights were present <300 m from the observation point. I collected observations on robin song during 18 April–8 June 2002 on 19 days with little or no wind or precipitation. I listened continuously for ≥20 min before the first robin sang, and recorded time of the first robin song in a circle of unlimited radius and time of chorus initiation, defined as when ≥2 robins first sang.
On 12 days I recorded song initiation times in Landingville, a rural town (pop. 175, 40°38′N, 76°07′W) 5 km east of the Schuylkill Haven site. A total of 22 outdoor lights were visible from the observation point, providing an intermediate level of artificial light.
I recorded song initiation times on 15 days at the Auburn Public Boat Launch (40°37′N, 76°07′W), an area dominated by farmland and a reservoir on the Schuylkill River, and thus with little artificial light. Only six outdoor lights were visible <1 km from the observation point. At this site I only recorded when the first robin sang because at most only 2–3 singing robins were detected on any given day and on some days only one singing robin was heard. The observation point was 2 km south of the Landingville site.
I usually began in Schuylkill Haven, proceeded to Landingville, then to the Auburn boat launch. Robin song initiation times differed enough among the three sites in mid-April that I considered it inefficient to sample just one site each morning. I heard only one pause in robin song >6.5 min before the morning chorus began on calm, dry days in 2002. Thus, I felt I did not miss the first robin song at the intermediate or low-light sites if I arrived >6.5 min before hearing a robin sing there.
On 13 April 2003, a night with clear skies, I measured light at each observation point. At two of the sites I also measured light at five randomly chosen locations within a 100 m radius. Light was measured with an Extech Instruments EasyviewTM 30 Light Meter with maximum resolution of 0.01 lux and a basic accuracy of ±3% of the reading (Extech Instruments, Waltham, MA). Minimum and maximum light readings were recorded during a 1 min period and then averaged.
Virginia Study Area
During April–July 2003 I recorded robin song initiation times in Lyon Park, Virginia, a densely populated residential suburb of Arlington, Virginia (38°53′N, 77°06′W), for comparison with observations made there in 1929 around the home of Harry A. Allard (Allard 1930). I located property owned by Harry A. Allard in Lyon Park during the 1920s by searching deed records in the Arlington County Courthouse. I arrived in Lyon Park >20 min before the first robin sang and made observations regardless of weather conditions using a circle of unlimited radius. I recorded wind conditions (windy or calm), temperature (°C), cloud cover (<50% or ≥50% cloudy), and precipitation (none, mist, or rain). Wind conditions initially were assigned based on whether local wind chimes were heard prior to song initiation. Daily wind measurements were also made upon arrival at the site during the last seven weeks of the field season with a hand-held KestrelTM 1000 wind meter (Nielsen-Kellerman, Boothwyn, PA). Throughout the study my definition of windy corresponded to an estimated wind speed >6 km hr−1. Once per weekend for five weeks in May and June 2003 I recorded when the robin chorus ended, defined as the first 7 min period during which <2 robins were heard singing.
After mid-April I measured light levels daily in Lyon Park during true night in the darkest nearby open area. Light was not measured during steady rain. I was primarily interested in effects of artificial light, rather than natural light, on song initiation time. As such, I felt that light measurements during true night would most accurately estimate the amount of artificial light present. If I then detected a correlation between these light levels and chorus initiation times I felt I would have some evidence that robin song behavior was being influenced by artificial light, or at least by noctural light. In contrast, light levels at the time of song initiation might reflect a mixture of artificial light and increasing natural light levels with the approach of dawn that could obscure a relationship between artificial light and singing behavior.
I extracted Allard's robin song initiation data from his figure 3 (Allard 1930:448) using dense tick marks published on both axes and a published grid overlaying the figure. I could not extract his weather data, and he did not measure light levels.
Maryland and District of Columbia Study Areas
Once per weekend for five weeks in May and June 2003 I recorded when robins began singing and when their chorus ended at a site with little artificial light at the USGS Patuxent Wildlife Research Center in Laurel, Maryland (39°03′N, 76°48′W). I arrived >20 min before the first robin sang.
Robins were heard singing at night near the National Mall in Washington, DC in 2003. The earliest singing robins I heard were along a fence around the White House. On 10 days during June–July 2003 I recorded light levels and robin singing behavior near this fence (38°54′N, 77°02′W), usually before proceeding to Lyon Park. At least 95 outdoor lights were visible from the White House fence observation point. I used the Patuxent site (low light) and the White House site (high light) as comparisons with Lyon Park (intermediate light).
Wisconsin Study Area
Aldo Leopold collected robin song initiation times in Madison, Wisconsin (43°04′N, 89°24′W) during March–July 1945 (Leopold and Eynon 1961). I obtained his data from the University of Wisconsin archives in Madison. Leopold recorded weather information and light levels daily, although his light meter could not measure light <0.13 lux.
Astronomical Data
Song initiation times were compared with the onset of local astronomical and civil twilight (U.S. Naval Observatory 2005). True night ends and astronomical twilight begins when the center of the sun is geometrically 18° below the horizon. Civil twilight begins in the morning when the center of the sun is geometrically 6° below the horizon. Moon phase data were obtained from program RedShift Version 3® (Maris Multimedia, Ltd. 1998). All times are local standard time (EST or CST).
Statistical Analyses
I modeled robin chorus initiation times relative to midnight at Schuylkill Haven and Landingville, and song initiation times relative to midnight at the Auburn boat launch by creating an a priori set of three linear regression models. The global model included day of year, site, and day of year*site. A second model included day of year and site; the third model included only a site effect. All analyses presented were conducted using Proc Mixed in SAS v. 8.2 (Littell et al. 1996), unless stated otherwise. All variables were treated as fixed effects in all analyses.
I created a priori sets of 26 competing linear regression models to model 1945 Madison song initiation times and 2003 Lyon Park chorus initiation times relative to civil twilight. For each site, one set of 13 models included day of year and day of year2, and models in the other set did not. The global model for Madison and the most general model for Lyon Park included day of year, day of year2, moon, clouds, moon*clouds, wind, rain, temperature and weekday. Null models included only day of year and day of year2 or only an intercept term. Four variables or groupings of variables were used to represent different hypotheses about what factors might influence robin song initiation time. I intended a priori for the variables “moon” and “clouds” and their interaction term “moon*clouds” to represent effects of natural light. The variable “light” was light level measured by my light meter and was intended to represent effects of artificial light in Lyon Park. The variable “light” did not appear in the Madison model set. The variables “wind,” “rain,” and “temperature” were selected to represent effects of natural sound or weather. I included the variable “weekday” to address possible effects of human-related noise or disturbance. One model was constructed to represent each of the four variable groupings, or hypotheses, except for the artificial light hypothesis in Madison. After constructing these models, the most general model, and the null model, I created 7–8 other models using different combinations of the above variable groupings. The model day of year, day of year2, light, and weekday was intended to represent the possibility that robin song initiation time was largely influenced by human-related factors in Lyon Park. Models with the variable “light” did not include the variables “moon” or “clouds” to minimize possible correlation among independent variables. As such, there was no global model for Lyon Park. I tested whether song initiation times were more variable in 2003 versus 1929 using a one-tailed variance ratio test (Zar 1999:139).
I compared robin chorus initiation times at Patuxent and Lyon Park in 2003 by constructing an a priori set of five linear regression models. The global model included day of year, site, and day of year*site. Three models included a subset of these variables. A null model included only an intercept term. This same model set was used to compare times at which robin choruses ended at Patuxent and Lyon Park in 2003.
Light level during true night in Lyon Park in 2003 was modeled with a set of 14 a priori linear regression models. The global model included day of year, day of year2, moon, clouds, moon*clouds, and precipitation. The variable “precipitation” = 1 if rain or mist was present; otherwise precipitation was zero. I created two identical sets of seven models, except that models in one set included day of year and day of year2 and models in the other set did not. The variables “day of year” and “day of year2” were included to address the possibility of a trend in nocturnal light during the field season. Null models included only the variables “day of year” and “day of year2” or an intercept term. The variables “moon” and “clouds” and their interaction term “moon*clouds” were included to represent natural light. Three models included different combinations of those three variables. The variables “clouds” and “precipitation” were paired in some models to represent possible effects of weather on light levels. In models of daily light level the variable “light” was log-transformed to achieve a normal distribution of errors.
Best models were selected using Akaike's information criterion corrected for small sample sizes (AICc; Burnham and Anderson 2002). The model with the lowest AICc is considered the best approximating model. The difference between the AICc of a given model and the AICc of the best approximating model (ΔAICc) can be used to determine the plausibility that a given model might be the best approximating model given the data. Models with a ΔAICc > 4 generally have little support. Akaike weights (wi) were calculated to evaluate support for each model and for individual variables.
I also analyzed song and chorus initiation times from all sites combined using linear regression models containing day of year, day of year2, site, and day of year*site. Contrasts were used with these two models to test for differences in song or chorus initiation times relative to civil twilight among sites that differed in levels of artificial nocturnal light (Cody and Smith 1997). Values reported are means ± SE except where indicated as SD.
Results
Pennsylvania Sites
On 16 April 2002 robins in Schuylkill Haven, PA were already singing by 01:11 EST, 155 min before the end of true night. Robin song previously has not been reported this early anywhere true night occurs. During the next eight weeks robins initiated their morning chorus during true night in Schuylkill Haven on 79% of all days sampled (Fig. 1, Table 1), and on average 116 min before onset of civil twilight. Choruses involved 10–20 robins at the sample site and numerous other robins nearby. I remained and listened to a given chorus up to 95 min. Light levels around my observation point averaged 3.91 lux (range = 0.11–11.79 lux, nlocations = 6) on 13 April 2003.
Table 1. Mean song and morning chorus initiation times of American Robins at several sites differing in levels of artificial nocturnal light, in min before local civil twilight. The sample size (n) is the number of days that song or chorus initiation times were recorded at each location and year
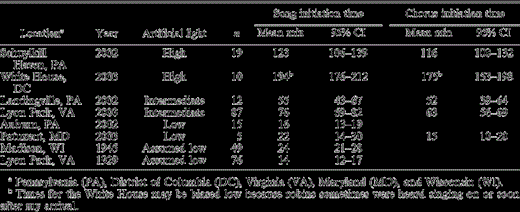
Table 1. Mean song and morning chorus initiation times of American Robins at several sites differing in levels of artificial nocturnal light, in min before local civil twilight. The sample size (n) is the number of days that song or chorus initiation times were recorded at each location and year
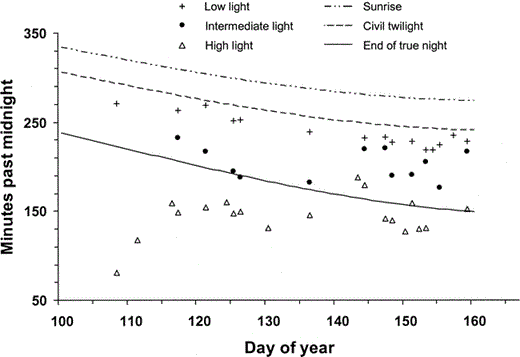
Figure 1. American Robin chorus initiation times at three sites in Schuylkill County, PA, May–June 2002.Choruses at a site with a high amount of artificial light (Schuylkill Haven) often began during true night, and appeared unrelated to sunrise. Song at a site with a low amount of artificial light (Auburn) began just before civil twilight, apparently in response to natural sunlight. Choruses at a site with an intermediate level of artificial light (Landingville) began at an intermediate time. Day of year 100 is 10 April; day of year 160 is 9 June
Robins began singing an average of 68 min later in Landingville, PA (Table 1, Fig. 1), a site with intermediate levels of artificial light (mean = 1.26 lux, range = 0.05–3.06 lux, nlocations = 6). There I began listening, on average, 26 min (range = 7–53 min) before the first robin sang. At the Auburn Public Boat Launch, a site with little artificial light (–0.03 lux, nlocations = 1), robins began singing near civil twilight, on average 39 min later than at Landingville, and at approximately the same time as indicated by previous studies in the early–mid 20th century (Allard 1930, Leopold and Eynon 1961, Table 1). I arrived at the Auburn site, on average, 29 min (range = 7–57 min) before the first robin sang. I heard a robin sing only once at the Landingville or Auburn sites before I had been present 6.5 min, and never before I had been present <6.0 min. From 18 April to 16 May (Day of year 108–136) I always arrived at each site >11.5 min before the first robin sang.
The global model was the best approximating model of song initiation times relative to midnight at the three Pennsylvania sites in 2002, and included a significant site*day of year interaction (Adj-R2 = 0.87). Parameter estimates in the best model were: β0= 378 ± 38; βDay of year = −0.99 ± 0.27; βSchuylkill Haven = −290 ± 50; βLandingville = −134 ± 61; βSchuylkill Haven*Day of year = 1.41 ± 0.36; βLandingville*Day of year = 0.70 ± 0.43 (Akaike weight for best model = 0.95, nmodels = 3).
Lyon Park, Virginia
Breeding robins were common to abundant in Lyon Park in 2003 and apparently so in 1929 (Allard 1930). Approximately 10–20 singing robins were present within my listening area in 2003 and numerous other robins sang nearby. A minimum of five different robins initiated daily song during the 2003 field season. During 7 April–16 July 1929 the first robin song in Lyon Park usually occurred just before civil twilight, and apparently never during true night (Allard 1930, Fig. 2, Table 1). In 2003, during the same time of year at the same location, robins initiated their morning chorus on average 49 min earlier, and during true night on 32% of all days sampled. On 76% of all sample days in 2003 robins initiated their morning chorus earlier (03:43 EST) than the earliest robin song ever recorded in Lyon Park during the entire 1929 breeding season. Song initiation times in Lyon Park were also more variable in 2003 than in 1929 (F86,78 = 8.7, P < 0.001).
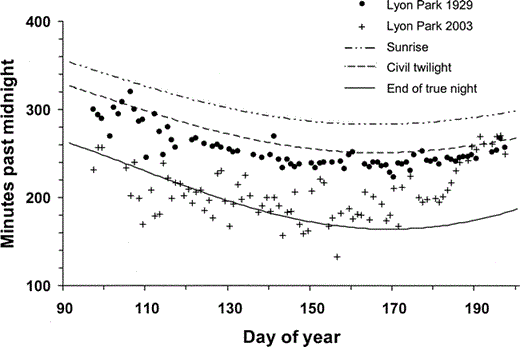
Figure 2. American Robin song initiation times in Lyon Park, Arlington, VA in 1929 and robin chorus initiation times at that same location in 2003.Choruses during 7 April–16 July 2003, began, on average, 63 min before onset of civil twilight, often during true night. In 1929 the first robin song occurred, on average, 49 min later, near onset of civil twilight. Day of year 90 is 31 March; day of year 190 is 9 July
Light levels near my observation point in Lyon Park in 2003 averaged 0.19 ± 0.15 SD lux during true night (range = 0.02–0.54 lux, ndays = 67), although large spatial heterogeneity in light levels was present. Light levels ranged from 6.8–72.5 lux on 21 April 2003 during true night at five random road-side locations in the brightest area ≤450 m from the observation point.
Daily robin choruses in Lyon Park in 2003 began earlier, relative to civil twilight, on bright nights and on weekdays (Table 2, best model Adj-R2 = 0.43, βlight = 47.6 ± 14.4; βweekday = 10.4 ± 5.1; ndays = 59, ΣAkaike weightlight = 0.88; ΣAkaike weightweekday = 0.69, nmodels = 26). The variables “moon” and “temperature” did not appear in any of the four best models. The brightest nights in Lyon Park in 2003 occurred with cloudy skies and misty conditions (best model: R2 = 0.74, β0 = −0.94 ± 0.14; βclear = −1.28 ± 0.15; βno precipitation = −0.86 ± 0.18; ndays = 56, Akaike weight for best model = 0.82, ΣAkaike weightclear = 1.0, ΣAkaike weightno precipitation = 1.0, nmodels = 14). No other model of light level had a ΔAICc ≤4, although all variables appeared in at least one model with an Akaike weight ≥0.09.
Table 2. Linear regression models of morning chorus initiation time for American Robins in Lyon Park, VA, 21 April–5 July 2003, ndays = 59, and in song initiation times in Madison, WI, 15 March–28 June 1945, ndays = 61. The dependent variable was minutes before onset of local civil twilight. “Day of year” goes from 1–365 in nonleap years, and was 74 for 15 March, 111 for 21 April, 179 for 28 June, and 186 for 5 July; “light” was a daily measure of light during true night in Lyon Park in 2003; “weekday” was coded one for Saturday and Sunday and zero for all other days of the week; “moon” was the proportion of the moon lighted; “clouds” was <50% or ≥50% cloud for Lyon Park, and “clear”, “partly cloudy”, or “cloudy” for Madison; “temperature” was °C; “wind” was “windy” or “calm,” and “rain” was presence or absence of rain. For the Madison data, clouds, wind, and rain were taken from a summary of Aldo Leopold's notes. The best three models for Lyon Park included level of nocturnal light with a positive slope, indicating that robins initiated morning choruses earlier relative to civil twilight on days when nocturnal light levels were higher. In contrast, the variable “light” appeared in only five of 23 models for Lyon Park with ΔAICc > 4. The global model for the Madison site had the most Akaike weight. Only the two (of 26) models for the Madison site that had a nonzero Akaike weight are shown. Deviance is a measure of model fit, K is the number of parameters in the model including the intercept and error term, AIC is an estimate of how well a model matches the unknown true mechanism that generated the data and AICc is the AIC adjusted for small sample size, ΔAICc is the difference between the AICc for a given model and the lowest AICc for the best approximating model, Akaike weights are estimates of relative support for each model and sum to one. Only models with ΔAICc < 4 are shown
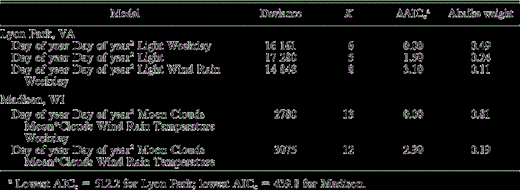
Table 2. Linear regression models of morning chorus initiation time for American Robins in Lyon Park, VA, 21 April–5 July 2003, ndays = 59, and in song initiation times in Madison, WI, 15 March–28 June 1945, ndays = 61. The dependent variable was minutes before onset of local civil twilight. “Day of year” goes from 1–365 in nonleap years, and was 74 for 15 March, 111 for 21 April, 179 for 28 June, and 186 for 5 July; “light” was a daily measure of light during true night in Lyon Park in 2003; “weekday” was coded one for Saturday and Sunday and zero for all other days of the week; “moon” was the proportion of the moon lighted; “clouds” was <50% or ≥50% cloud for Lyon Park, and “clear”, “partly cloudy”, or “cloudy” for Madison; “temperature” was °C; “wind” was “windy” or “calm,” and “rain” was presence or absence of rain. For the Madison data, clouds, wind, and rain were taken from a summary of Aldo Leopold's notes. The best three models for Lyon Park included level of nocturnal light with a positive slope, indicating that robins initiated morning choruses earlier relative to civil twilight on days when nocturnal light levels were higher. In contrast, the variable “light” appeared in only five of 23 models for Lyon Park with ΔAICc > 4. The global model for the Madison site had the most Akaike weight. Only the two (of 26) models for the Madison site that had a nonzero Akaike weight are shown. Deviance is a measure of model fit, K is the number of parameters in the model including the intercept and error term, AIC is an estimate of how well a model matches the unknown true mechanism that generated the data and AICc is the AIC adjusted for small sample size, ΔAICc is the difference between the AICc for a given model and the lowest AICc for the best approximating model, Akaike weights are estimates of relative support for each model and sum to one. Only models with ΔAICc < 4 are shown
Maryland, Wisconsin, and District of Columbia Sites
Robin choruses in Lyon Park in 2003 began 33 min earlier than robin choruses on adjacent days that same year at a Patuxent site with virtually no artificial light (nPatuxent days = 5, nVA days = 10, P < 0.001, Akaike weight for model with site effect only = 0.86, nmodels = 5, 1 site visited per day; light rangePatuxent = 0.01–0.09 lux, ndays = 3). Timing of the end of the morning robin chorus did not differ between Lyon Park and Patuxent in 2003 (nPatuxent days = 5, nVA days = 5, Akaike weight for intercept only = 0.47; Akaike weight for site effect model = 0.45 with chorus ending 8 min earlier at Patuxent, nmodels = 5).
During 7 April–28 June 1945 robins in Madison, Wisconsin began singing on average 24 min before civil twilight and did not sing during true night (Leopold and Eynon 1961, Table 1). The global model was the best approximating model of robin song initiation time at this site (Table 2). Slope estimates in the global model included βno wind = 8.7 ± 2.1, βtemperature = 1.6 ± 0.3, βno rain = 10.7 ± 3.2, βweekday = 6.4 ± 2.5, βmoon = 14.4 ± 9.0, βclear = 0.4 ± 6.8, βcloudy = 0.8 ± 6.3, βmoon*clear = 3.4 ± 10.1, and βmoon*cloudy = −14.6 ± 10.1. Published light levels in this study during March–June 1945–1948, were generally 0.13–0.44 lux (ndays = 106) when the first robin sang (Leopold and Eynon 1961). Whether light levels were lower during true night is unknown.
Robin song began approximately 18 min earlier on clear, apparently moonlit, mornings than on cloudy mornings in Madison in 1945, based on expected values from the best model of daily song initiation time (best model: R2 = 0.76, ndays = 61, Leopold and Eynon 1961, Table 2). However, robin choruses began earlier on cloudy nights than on clear nights in Lyon Park in 2003 (fourth best model: βNo Clouds = −11.5 ± 5.1).
The seemingly brightest area I sampled in 2003 was the White House in Washington, DC (mean = 1.06 lux during true night at the darkest accessible nearby open area, range = 0.94–1.18 lux, ndays = 9, nlocations = 1). The earliest I heard a robin sing at this site was 255 min before civil twilight on 10 May 2003. Robin morning choruses (nrobins present ≥10, nrobins singing ≥2–3) began there before 02:20 EST on 10 of 10 days sampled during 20 June–14 July 2003. On five days I stayed and listened to these choruses for >20 min, and on one day I stayed and listened to a chorus >40 min.
Multisite Analysis
I created contrasts based on the site groupings in Table 1. Contrasts indicated robin song initiation times in Lyon Park differed between 1929 and 2003 (P < 0.001), between sites with high levels of artificial light versus sites with an intermediate level of artificial light (P = 0.02), and between sites with intermediate versus low levels of artificial light (P < 0.005). Song initiation times did not differ between sites with known little versus assumed little artificial light (P = 0.34).
Contrasts for chorus initiation times detected a difference between sites with high versus intermediate levels of artificial nocturnal light (P < 0.005), and a weak difference between Patuxent, a low light site, and two sites with an intermediate amount of artificial nocturnal light (P = 0.10).
Discussion
All results in this study suggest that robins initiate their morning chorus earlier in areas with high levels of artificial light, and that robins in areas with little artificial light continue to initiate song at approximately the same time, relative to civil twilight, as did robins historically (Allard 1930, Leopold and Eynon 1961, Fisler 1962). Cloud cover and mist appear to amplify the effect of artificial light by trapping that light near the ground in urban areas, making the night even brighter and prompting robins to initiate song even earlier.
Since I usually sampled more than one of the three Pennsylvania sites in the same morning, a significant site effect is trivial in a statistical sense when modeling song initiation times among these areas. However, I suggest the difference in slopes among sites is not an artifact of the sampling approach, but rather is evidence that artificial light now plays a larger role than natural light in song initiation by urban American Robins. Song initiation times at the brightly lit Schuylkill Haven site did not appear to be influenced by onset of civil twilight, but song initiation times appeared to track civil twilight at the study area with little artificial light.
I assume light levels were lower in Lyon Park in 1929 than in 2003. Streetlights now number 81 000–100 000 in nearby Washington, DC (J. Abdi, DC Department of Public Works, pers. comm.; M. Dorsey, DC Department of Public Works, pers. comm.). Gas streetlights apparently numbered <12 500 during their peak use in Washington, DC in the 1920s (Williams 2002), although they were then being replaced with an unknown number of electric street lights. Also, in 2003 there appeared to be several homes on Allard's former property, suggesting the density of the human population in the surrounding area may have increased since 1929. The human population of Arlington County, Virginia increased from 26 700 in 1930 to 189 000 in 2000 (U.S. Census Bureau 2005). Allard (1930:459–460) was skeptical that existing street lights affected the singing behavior of birds. However, he did not rule out the possibility that artificial light of the “proper intensity and quality” (p. 460) might have an affect on singing behavior. I suggest that as the human population in Arlington County has increased, so has the level of artificial light in the region, and that artificial nocturnal light has now reached a level at which it strongly affects robin singing behavior.
Measured light level also could not be included in the 1945 models for Madison, Wisconsin, primarily because the light meter used then could not measure light levels <0.13 lux. However, comparison of variables included in the best 2003 model for Lyon Park with those in the best 1945 model for Madison suggests that perhaps artificial light has become a dominant stimulus in initiating song in urban robins.
All results presented here are consistent with the hypothesis that light initiates robin song, and with the hypothesis that light pollution is affecting robin song behavior at a population level. Nevertheless, these results are correlational, and as such can provide only relatively weak inference (Romesburg 1981). A variable other than light level potentially could be affecting robin singing behavior. For example, loud noise appears to increase alertness and avoidance behaviors of some bird species (Ward et al. 1999). If high noise levels or human disturbance were responsible for awakening robins and prompting them to sing during true night in Lyon Park in 2003 I would have expected robin song to begin there earlier on weekends, since human activity between 01:00 and 03:00 EST was greater in the area on weekends. My results suggested that robins in Lyon Park began singing later on weekends. Robins also sang later on weekends in Madison in 1945, although I do not know why. I also did not address the possibility that differences in food availability among sites might influence song initiation times of American Robins. A suburban population of Florida Scrub-Jays (Aphelocoma coerulescens) initiated breeding activity earlier in the year than did a nearby population in rural habitat (Schoech and Bowman 2003), and the authors suggested that suburban scrub-jays could breed sooner because of an increased availability of high-quality food sources in urbanized areas.
The abundance of robins within a site might also affect how vigorously they sing (Leopold and Eynon 1961). Robins appeared to be fairly abundant at all sites with at least an intermediate amount of artificial light, although there were seemingly fewer robins in Landingville than in Schuylkill Haven. I do not know whether abundance of robins changed within a site during a field season. Perhaps some migrants were still present in April. The earliest that a robin ever sang within a given field season relative to civil twilight occurred between late March and mid-April in Lyon Park in both 1929 (Allard 1930) and 2003, in Michigan (Fisler 1962), in Wisconsin (Leopold and Eynon 1961), and in Schuylkill Haven. Perhaps such early singing behavior at that time of year is an attempt to establish or protect territories while abundance of adult robins is particularly high due to the presence of transient individuals.
Manipulative experiments enable the strongest inference (Romesburg 1981). Light levels could be randomly manipulated nightly at several rural parks or athletic fields equipped with electric lights, and robin song initiation times could be monitored in conjunction with levels of light, sound, and perhaps food resources and gonadal hormone levels. Light pollution might affect physiology and endocrinology of birds is several ways. Artificial light may increase levels of testosterone or its active androgenic and estrogenic metabolites thereby causing male robins to awaken earlier and sing for longer periods each day (Van Duyse et al. 2005). Artificial light may also cause neurological changes in birds, for example in the higher vocal center, thereby potentially affecting song behavior (Dawson et al. 2001). Song initiation times might not be limited by availability of light after the amount of nocturnal light reaches a certain high level, or else robins in areas with a large amount of artificial light might be expected to sing continuously from dusk to dawn. Perhaps robins in such areas simply require a minimum threshold of sleep before they can begin singing.
Other potential future studies, with or without a manipulative component, might address possible effects of light pollution on robin population dynamics. Light pollution appeared to make robins sing longer each morning in the present study. As such, robin survival in brightly lit areas might be reduced if energy demands or predation risk are elevated there (Kacelnik and Krebs 1982, McNamara et al. 1987, Hutchinson et al. 1993). Alternatively, light pollution might increase robin productivity if robins can forage or seek mates during longer periods each day.
Acknowledgments
I thank K. Knight for encouraging me to study formally my casual observations of robin morning song, and T. Clauser, W. Manning, B. Schermetzler, and various law enforcement personnel for logistical support. J. Nichols, B. Kendall, A. Royale (Patuxent Wildlife Research Center, Laurel, MD), D. Dobkin, T. Sachtleben, and an anonymous referee provided valuable advice on earlier versions of the manuscript. I especially acknowledge H. A. Allard and A. Leopold whose data I have incorporated into this study. This paper is dedicated to M. Gordon.
Literature Cited

{kind=link}
{kind=link}