Microbial Ecology of the Planetary Boundary Layer


1
Institut des Géosciences de l’Environnement, Université Grenoble Alpes, CNRS, IRD, Grenoble INP, Grenoble, 38400 Saint-Martin-d’Hères, France
2
Environmental Microbial Genomics, CNRS UMR 5005 Laboratoire Ampère, École Centrale de Lyon, Université de Lyon, Écully, 69007 Lyon, France
*
Author to whom correspondence should be addressed.
Atmosphere 2020, 11(12), 1296; https://doi.org/10.3390/atmos11121296
Submission received: 17 October 2020
/
Revised: 28 November 2020
/
Accepted: 28 November 2020
/
Published: 30 November 2020
(This article belongs to the Section Aerosols)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Aerobiology is a growing research area that covers the study of aerosols with a biological origin from the air that surrounds us to space through the different atmospheric layers. Bioaerosols have captured a growing importance in atmospheric process-related fields such as meteorology and atmospheric chemistry. The potential dissemination of pathogens and allergens through the air has raised public health concern and has highlighted the need for a better prediction of airborne microbial composition and dynamics. In this review, we focused on the sources and processes that most likely determine microbial community composition and dynamics in the air that directly surrounds us, the planetary boundary layer. Planetary boundary layer microbial communities are a mix of microbial cells that likely originate mainly from local source ecosystems (as opposed to distant sources). The adverse atmospheric conditions (i.e., UV radiation, desiccation, presence of radicals, etc.) might influence microbial survival and lead to the physical selection of the most resistant cells during aerosolization and/or aerial transport. Future work should further investigate how atmospheric chemicals and physics influence microbial survival and adaptation in order to be able to model the composition of planetary boundary layer microbial communities based on the surrounding landscapes and meteorology.
1. Introduction
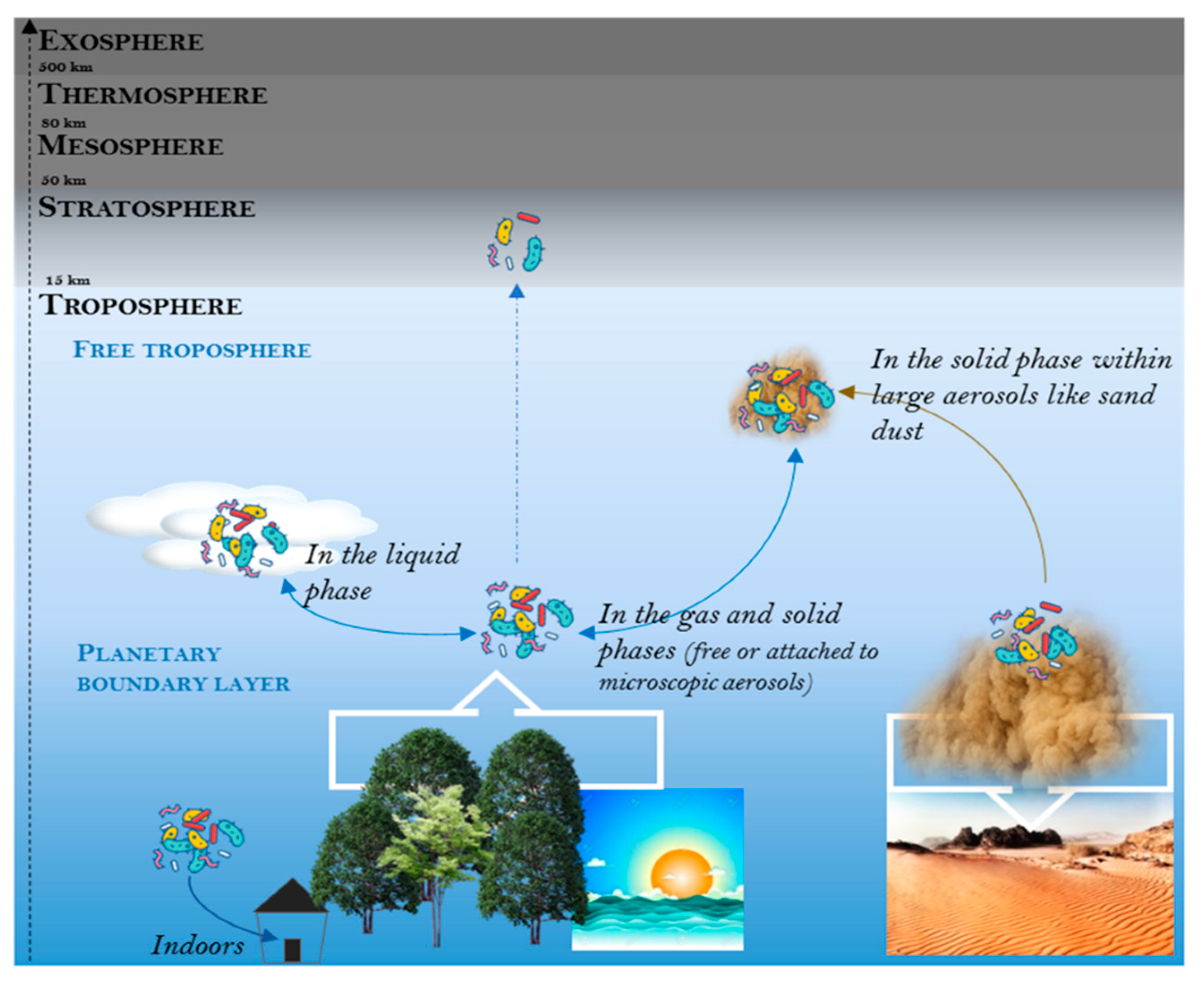
Aerobiology is a growing research area that covers the study of aerosols of a biological origin (i.e., bioaerosols) suspended in the atmosphere, from the air that directly surrounds us (both indoors and outdoors) to space by going through the different atmospheric layers (Figure 1). Bioaerosols include plant debris, pollen, microorganisms (bacteria, fungi, viruses, protozoans, etc.) as well as biological secretions [1,2] which are mainly emitted by natural (forests, oceans, deserts, etc.) and urbanized Earth surfaces (agricultural fields, waste water treatment plants, cities, etc.) at different emission rates [3,4]. Airborne microorganisms, especially bacteria, archaea, and fungi, are of particular interest as they represent living and potentially metabolically active cells light enough to be lifted high in the atmosphere by upward airflow [5,6,7]. During extreme meteorological events such as volcano eruptions and dust storms, sand-dust associated microorganisms can be ejected tens of kilometers high in the atmosphere before landing back on the Earth’s surface thousands of kilometers away [8,9]. Microorganisms from the Bacillus and Micrococcus genera are commonly recovered from the stratosphere [6,10,11]. Research on the “high life” [5] intends to evaluate the global dispersion of microorganisms around our planet as well as establish the upper limit of the biosphere boundary and infer the probability of life in the universe [12,13]. Other interests are driving investigations of microorganisms in the lowest atmospheric layer, the troposphere, that surrounds the Earth’s surface. The role of airborne microorganisms in meteorological processes such as cloud formation and precipitation [14,15,16,17], atmospheric chemistry [14,18,19,20,21], and air quality [9,22,23,24] are currently motivating multidisciplinary investigations on microbial communities in the troposphere. The capacity of microorganisms to be transported through the air has raised concern about the role airborne microorganisms might play in public health with the potential dissemination of plant and human pathogens as well as allergens [9,25]. The importance of airborne micro-organisms should motivate the inclusion of a biological component into the existing and future observing infrastructure of the earth atmosphere.
Subject to gravity, aerosols (or particulate matter) as well as bioaerosols become concentrated in the lower part of the troposphere that is called the planetary boundary layer (Figure 1). Microbial concentrations thus usually show a vertical stratification from the bottom to the top of the troposphere with average estimated bacterial concentrations of 9 × 102 − 2 × 107 cells/m3 in the planetary boundary layer (based on six qPCR-based studies: [26,27,28,29,30,31] and 4 × 101 – 8 × 104 cells/m3 in the highest part of the troposphere called the free-troposphere (based on three qPCR-based studies [32,33,34]). Yet, microbial concentration estimations vary between investigations, which are based on different sampling strategies. Heterogeneity of methodology in aeromicrobiology is a current issue that will be discussed in a subsequent section. The troposphere is the most dynamic layer in terms of chemistry and physics of aerosols and harbors complex chemical reactions and meteorological phenomena that lead to the coexistence of a gas phase, liquid phases (i.e., cloud, rain, and fog water) and solid phases (i.e., microscopic particulate matter, sand dust) (Figure 1). The various atmospheric phases represent multiple biological niches that might harbor different microbial communities (Figure 2) and different microbial concentrations that might be due to significant differences in physico-chemical characteristics that constrain either microbial life within the niches or the destruction of specific members from their source. Liquid-phase associated microbial communities (i.e., associated to cloud, rain, and fog water) might be particularly different from the microbial communities of the dry phase of the troposphere [35] (Figure 2). Reviews and field investigations that are specifically related to cloud-associated microorganisms can be found in [14,36,37,38,39]. This review aims to synthesize knowledge and gaps of knowledge regarding microbial communities in the dry phase of the troposphere (i.e., gas and solid phases). Available data allow for assumptions regarding what controls microbial communities in the troposphere, especially the planetary boundary layer.
2. Microbial Cell Dynamics in the Troposphere
While airborne microbial concentrations have been repeatedly measured in the lower troposphere around our planet using culture-dependent and molecular analyses [27,28,29,30,31,42,43,44], cell concentrations in the upper troposphere and stratosphere remain unknown. In-flight collection of microorganisms in the upper troposphere and stratosphere remains expensive and an engineering challenge which, due to the likely very low cell concentrations in the stratosphere, suffers from aircraft-associated contamination [6,7,13]. It is likely that airborne microbial concentrations tend to decrease from the lower troposphere up to the upper troposphere [45] (up to fifteen kilometers) and the stratosphere [32,46] although the rate of decline is unknown. The vertical gradient in microbial concentration suggests that microbial cell fluxes might be upward in the atmosphere. Downward microbial cell flow coming from the stratosphere and the high troposphere might be quantitatively minor (Figure 3) and thus have little impact on airborne microbial concentrations and the composition of the low troposphere. Exceptions to the rule might occur during extreme meteorological events such as volcano eruptions, hurricanes, and sand dust storms. In the latter case, microorganisms associated to large particulate matter, such as macroscopic sand dust, could be lifted high in the troposphere, travel along global air masses over thousands of kilometers then settle back to the Earth’s surface due to gravity, precipitation, and atmospheric circulation [9,12,13,47]. The downward flow of large diameter-particles and their associated microorganisms that have been transported through the upper troposphere might disturb the structure of airborne microbial communities of the downwind area both quantitatively, with an increase in microbial concentration, and qualitatively, with a change in microbial composition and size distribution [9,31,47,48,49,50,51]. Airborne microbial cells exist mainly as aggregates or attached to particulate matter (size range from less than one nanometer up to hundreds of micrometers like sand dust), while airborne fungi exist mainly as single spores [52]. Microbial cells entering freely in the atmosphere can attach to existing particulate matter or other microbial cells [53]. Conversely, particle-attached microbial cells can detach from their support in the air. Based on a compilation of data from more than one hundred investigations, Clauss et al. [52] determined that 15% of cultivable airborne bacterial cells were on particles <2.1 µm (size) and 25% on particles >7.2 µm, and that cultivable airborne fungal spores and cells were mainly distributed on particles between 1 and 3.2 µm (median-based values) on average in outdoor air. The size distribution was shown to depend on the aerosolization processes (sea spray, bubble-bursting in whitecaps, blasting and splashing raindrops, etc., [54,55,56]) and meteorological conditions at the time of aerosolization such as air relative humidity [52]. Particulate matter and bioaerosols in the planetary boundary layer, the lower part of the troposphere (Figure 1), are moved to the rhythm of the air masses that are subject to mechanical and thermal convective turbulence partly controlled by the ground roughness and the Earth’s surface heat [57]. As a consequence, the planetary boundary layer’s height changes according to location and even time of day throughout the year. In contrast, the free troposphere (above the planetary boundary layer) tends to be vertically stable and driven horizontally by geostrophic wind. Aerosolized microbial cells generally enter the planetary boundary layer from which a yet unknown quantity might be transferred to the free troposphere that might depend on the meteorological conditions and global air circulation. Within the planetary boundary layer, airborne microorganisms might have a residence time of a few days before returning to the Earth’s surface due to gravity or precipitation (model assuming that microbial cells behave like non biological aerosols, [58]). In the free troposphere, their residence time might be several days during which they might be transported over long distances [12]. Despite an obvious continuum of the troposphere and because of differences in aerosol dynamics, chemical composition, and physical conditions, investigations on the vertical distribution of airborne microbial communities showed different microbial communities in the planetary boundary layer and free troposphere [32,46] and suggested that some microbial taxa might be filtered out during vertical transport [46]. Due to their size, the largest and densest airborne microbial cells might be less prone to reaching the free troposphere than lighter cells. This hypothesis was supported by the observed increase in the ratio between bacteria and fungi at a remote mountain site in Austria (3106 m above sea level, [46]). Another explanation could be that microbial cells floating in the free troposphere have more time to undergo selection and adaptation to the abiotic conditions as compared to those in the planetary boundary layer, so that only the microorganisms that are the most resistant to the harsh tropospheric conditions (UV radiation, cold temperature, radicals, etc.) survive (physical selection and microbial adaptation will be discussed in the following sections). Thermophilic strains with high resistance towards extreme conditions, which are often identified in heavy dust events, were shown to be ubiquitous and significantly increased in relative abundance in the free troposphere as compared to the planetary boundary layer at a remote mountain site in Austria (3106 m above sea level, [46]).
3. Structuring Factors of Microbial Communities in the Planetary Boundary Layer
3.1. Surfaces, Aerosolization, Local Versus Distant Sources
Airborne microorganisms originate mainly from both natural (forests, oceans, deserts, etc.) and urbanized surfaces (agricultural fields, waste water treatment plants, cities, etc.). Burrows et al. [3,4] constrained a general atmospheric circulation model using data from the literature and estimated that 1024 bacteria are emitted into the atmosphere each year at a global scale. Observations of the microbial diversity in the planetary boundary layer showed that airborne microorganisms from one air sample might come from many different ecosystems (plants, soil, ocean, etc.) that might explain the observed large taxonomic diversity of airborne microbial communities. Aerosolization from Earth surfaces depends mainly on the landscapes (forest, grassland, ocean, etc.) as well as the current meteorological conditions [59]. Oceanic surfaces were shown to emit less than terrestrial surfaces [4]. Among terrestrial surfaces, grasslands might be the most effective emitters of microorganisms, while ice potentially emits 100 times fewer microbial cells [4]. Only a few studies investigated the selective propriety of aerosolization mechanisms, yet aerosolization might be the first critical process controlling which microorganisms are present in the air [60,61]. In acting as a filtering process, aerosolization might mediate the ratio between bacterial and fungal cells observed in the air as well as influence the ratio between the different populations within these kingdoms. Microscopic and molecular biology analyses showed that bacterial cells are generally in higher concentration compared to fungal cells (i.e., hyphal fragments) and spores in the planetary boundary layer [32,44,46,48,62,63]. Small-sized cells, such as bacterial cells that are usually 10 times smaller in size than fungal cells, might be preferentially aerosolized. Womack et al. [64] observed that the Amazonian forest air was more loaded in Ascomycota than Basidiomycota fungi. They suggested that Ascomycota fungi might be preferentially aerosolized because of their single-celled and filamentous vegetative growth forms that are much lighter than Basidiomycota spores. Low wind speed might be more effective in lifting light cells and light particulate matter while strong wind speed might also lift macroscopic dust and associated microorganisms. Aerosolization could also depend on the physiological properties of the cell membrane. Specific bacterial taxa (e.g., Actinobacteria and some Gammaproteobacteria) and lipid-enveloped viruses have been proposed to be preferentially aerosolized from oceans as a result of hydrophobic properties of their cell envelope [60]. At a given site, aerosolization specificity (both passive and active) might depend on the current meteorology and surface conditioning (vegetation height, presence of a snow cover, soil composition, etc.) (see [59,64,65,66], and [67] for fungi). On-site studies showed that the ratio between fungal fragments (hyphae fragments) and spores from soil might be of one, and tightly dependent on meteorological conditions (wind speed and direction) as well as fungal species [68]. Wind turbulence, blasting, and splashing raindrops might mediate the introduction of microbial cells into the air. Over oceanic surfaces specifically, sea spray and bubble-bursting in whitecaps (foam crest over the waves) and breaking waves are critical processes affecting the emission of microbial cells to the atmosphere [54,55,56]. It is not yet clear in which conditions rain droplets contribute either to aerosolization or washout of microbial cells from the air [69], but surface temperature, composition, and relative humidity as well as rain intensity seem to play important roles. Recent data showed that, of the meteorological factors, wind, temperature, and relative humidity are particularly important in establishing the planetary boundary layer microbial community composition [27,59,70] (Figure 3). They can mediate the different inputs of microbial cells from the different surrounding surfaces (for example with a change in wind direction), and their role would be even more important if the surrounding landscapes showed a high diversity (high richness and low evenness) in ecosystems [59]. Sites characterized by strong changes in meteorological conditions over time would show an even larger temporal variability in the composition of planetary boundary layer microbial communities [59]. Meteorology is also responsible for the transformation of the surface conditioning (vegetation height, presence of a snow cover, etc.) throughout the year. A change in the surface conditioning might lead to a change in the Earth’s surface microbial communities and thus a change in the diversity of the aerosolized microbial cells observed throughout the seasons [42,71,72,73,74]. Large and visible particles such as sand dust have provided evidence for long-range transport of aerosols (i.e., transport over hundreds of kilometers) including bioaerosols. The aerial long-range transport of microorganisms was particularly studied during dust storm events originating from Asian and African deserts [8,9,12,31,47,49,50,75,76,77,78,79,80,81,82,83,84,85]. Powerful natural events like dust storms, hurricanes, volcano eruptions, and forest fires can lift microorganisms up into the high troposphere and stratosphere, and lead to the aerial transport of microorganisms far away from their source environments [9]. Some studies observed a complete change of the tropospheric microbial community abundance and structure of the downwind sites [31,47,48,49,50,51,86]. Dust-associated microbial communities seem to be taxonomically different from the ones of the troposphere (culture-based approach) and in higher concentration (up to 10 times higher, see [31,47,48,51]). Under milder weather, wind speed might mediate the inputs of microbial cells coming from distant sources. Collected airborne microbial communities might be a mix of microbes originating from local sources and distant sources whose relative contribution might depend on the global meteorological characteristics (i.e., influence of the free-troposphere, high wind speed) and the geographical site (i.e., altitude from sea level, surface conditioning). Recent investigations [59,73] including a global-scale investigation [59] strongly suggested that proximity has likely a larger impact and that local sources (e.g., surrounding landscapes) are the main contributors to the airborne microbial community composition (Figure 3). During aerial transport, airborne microorganisms would be deposited through precipitation and gravitational settling. The modeled residence time of airborne microbial cells defined as a round and free aerosol of 1 µm of diameter was estimated to be 3.4 days on average [3]. Still, free airborne microbial cells of 1 µm might represent a small fraction (<15%, see [52]) of the planetary boundary layer microorganisms that undergo the physical selection that might occur during aerial transport (as discussed below).
3.2. Physical and Chemical Conditions that Might Constrain Microbial Life in the Planetary Boundary Layer
Metagenomic investigations of the complex microbial communities of many ecosystems have provided evidence that microorganism functional signatures reflect the abiotic conditions of their environment [87,88,89,90]. In the planetary boundary layer, like in any atmospheric layer, physical conditions and a variety of chemical substances interacting with airborne microorganisms might have an effect on them. Specifically, UV radiation, temperature shocks, desiccation, as well as the presence of free radicals might constrain microbial life in the dry phase of the planetary boundary layer. These conditions might be controlling factors in leading to the survival and/or development of microbial taxa with specific resistance mechanisms and/or functions in the atmosphere. On the one hand, the physical and chemical conditions might cause the death of non-resistant cells, a process we consider as a physical selection. Surviving resistant cells might develop if they are active and growing while undergoing a microbial adaptation (i.e., genetic changes in the genome in response to the physical and chemical conditions) to the atmospheric environment.
3.2.1. Physical and Chemical Conditions Characterizing the Atmosphere
The highly energetic wavelengths of UV radiation (UV-C ~190–290 nm and UV-B ~290–320 nm) might be responsible for direct DNA damage that could be lethal. Longer wavelengths (UV-A ~320–400 nm and visible light ~400–800 nm) contribute to intra-cellular reactive oxygen species (ROS) production that can cause subsequent oxidative damage to DNA, RNA, lipids, and proteins, altering microbial metabolism and survival [91,92]. Data on the impact of UV radiation on airborne microorganisms come mainly from investigations using high UV levels such as those found in the upper troposphere or stratosphere [5,93,94,95,96], and might not apply to the planetary boundary layer. The stratosphere supports by far the highest levels of UV radiation found on Earth, as levels increase by around 11% with every 1000 m in altitude (WHO). Smith et al. [94] showed that UV radiation was the most biocidal factor in the low stratosphere and could kill up to 99.9% of Bacillus subtilis spores after 96 h. However, the authors pointed out that spore resistance might be dependent on the environment that the cells germinated [97,98,99], and consequently, UV resistance might have been higher if the spores were directly isolated from the stratosphere and not germinated in culture media as was done in the study. Microbial strains isolated from the upper troposphere and lower stratosphere exhibited a higher resistance to UV radiation as compared to those from the planetary boundary layer [95]. Some Deinococcus and Streptomyces strains showed an extreme UV resistance and tended to form aggregates in culture medium. These aggregates were suggested to be a protection mechanism [95]. With the exception of sporulation and cell aggregation, no other protective mechanisms against UV radiation have been observed in airborne microbial communities. Although UV radiation levels can be extremely high and destructive in the upper atmosphere, it may not be the case in the planetary boundary layer. Moreover, UV levels perceived by planetary boundary layer microorganisms might differ depending on geography (for example the tropics harbor higher UV levels) and surface conditioning (i.e., surface reflectance) [100].
In the same way, air temperature is highly dependent on the latitude and longitude at a given altitude, and decreases by 0.6 to 1 °C for every 100 m increase of altitude. Within the planetary boundary layer, upward aerial transport of microorganisms with high-speed winds could occur rapidly and airborne microorganisms might suffer large temperature shocks. Airborne microorganisms present in an air parcel transported from the surface to a 1 km altitude can undergo a temperature decrease of 5 to 10 °C and a substantial increase in relative humidity [57]. Cold temperatures and freeze-thaw cycles generally occur at high latitudes and high altitudes. In other ecosystems, they have been shown to slow down microbial metabolism, decrease membrane fluidity, and influence protein refolding. Freeze-thaw cycles could additionally lead to mechanical stress that might damage the cell membrane [101,102,103]. Freeze-thaw cycles were shown to alter the survival of microbial strains following UV radiation, H2O2 exposure, and osmotic shock when these factors were tested individually on strains isolated from clouds belonging to Pseudomonas, Sphingomonas, Arthrobacter, and the yeast Dioszegia [104]. To date, the role that temperature shocks and freeze-thaw cycles might play on microorganisms in the planetary boundary layer remains largely unexplored. Their impact might be closely related to air relative humidity (RH). A large range of RH values could be found within the planetary boundary layer depending on latitude, altitude, and time of day. Investigations on the survival of aerosolized microorganisms under different RH showed different results depending on the species [105,106]. While the survival of airborne Flavobacterium was not affected by RH ranging from 25 to 99% at 24 °C [105], mid-range RH negatively impacted mycoplasma survival, but not RH values outside of this range [106]. Desiccation, like radiation, tends to induce DNA damage [107,108]. In the environment, desiccation resistance is generally associated to ionizing radiation resistance [107,109,110,111,112], yet the nature of the underlying mechanisms remains unknown. Changes in RH are linked to evaporation/condensation cycles of water vapor. In a water droplet, evaporation can concentrate metabolites in the near environment of the cells by up to 1000 times [104]. Evaporation/condensation cycles induce osmotic changes, leading to water fluxes between the intracellular and extracellular compartment of the cell to maintain osmolarity. These water fluxes can provoke cell damage, increase the concentration of metabolites in cells, and increase the concentration of compounds like radicals and metals around the cell [113,114]. Alsved et al. [115] showed that during evaporation, Pseudomonas syringae survival was enhanced when the relative humidity rapidly reached the level where salts become solid. Hence, small and salty liquid droplets were suggested as a more suitable environment when exposed to evaporation than large and slightly salty liquid droplets [115]. Still, the effect of evaporation/condensation cycles on airborne microbial communities and the mechanisms they use to protect themselves remain largely unknown.
The potential impact of the oxidizing nature of the atmosphere that is characterized by an enhanced presence of radicals (OH, O2−), nitrate radicals, and OH precursors such as hydrogen peroxide (H2O2) [57,116] on airborne microorganisms has been mainly investigated in cloud water. Joly et al. [104] tested the effect of different concentrations of hydrogen peroxide on the survival of different microbial strains isolated from cloud water. They showed that the 50% lethal concentration of H2O2 was different among the strains, and 10 times higher than the typical concentration found in puy de Dôme (France) cloud water. Increases in ROS could occur during other environmental stresses like UV radiation as discussed above.
3.2.2. Effects of Atmospheric Conditions on Microbial Life during Aerosolization and Aerial Transport
The atmospheric physical and chemical conditions described above might impact microorganisms during aerosolization and aerial transport. Laboratory studies on E. coli showed that up to half, and even 99%, depending on the aerosolization mechanism and intensity, of the aerosolized bacterial cells suffered from cell membrane damage after 10 min of aerosolization [117], which might lead to subsequent death. Another study showed that the viability of Pseudomonas fluorescens bacteria decreased by over 50% after 90 min of continuous aerosolization [118]. The sublethal damages occurring during aerosolization were associated to a differential gene expression of respiratory, cold-shock, metabolism, and more generally stress-response genes [117,118,119,120,121]. Aerosolization in small and salty liquid droplets that dry rapidly was shown to induce a higher survival rate of Pseudomonas syringae [115]. Since these kinds of liquid droplets are mainly formed from liquid environments like oceans, aerosolization from oceans might promote the survival of cells as compared to dry environments like soil [115]. Airborne microbial communities might be a subsample of the surface microbial communities that underwent a selective process during both aerosolization and aerial transport. Microorganisms harboring specific physical characteristics (spore, membrane characteristics, aggregation of cells, etc.) and/or genetic and enzymatic microbial properties (efficient DNA repair mechanisms, etc.) might be more likely to survive the conditions (UV radiation, desiccation, etc.) encountered post aerosolization. While microbial cells able to resist to these conditions have been observed in the air, the question about whether these resistant cells represent the majority of the airborne microbial community remains. Little is known about the survival mechanisms of both airborne bacterial and fungal cells and the ratio between resistant and sensitive cells in the air. Survival mechanisms such as dormancy, sporulation, aggregation among cells, or with particulate matter as well as specific microbial resistance to the extreme conditions encountered in the atmosphere are relatively common in the environment [104]. Fungal spores have evolved to survive and disseminate through the planetary boundary layer and are known to be particularly resistant to atmospheric conditions and especially to desiccation, UV radiation, and oxidative stress [122]. Yet, their resistance might have been selected for on Earth surfaces before being aerosolized. A recent comparative metagenomic study showed that fungi were more dominant relative to bacteria in air compared to the other planetary bound ecosystems (soil, ocean, etc.) (Tignat-Perrier et al., in revision). This suggests a selective process for fungi during aerosolization and/or aerial transport and that fungi might likely survive aerosolization and/or aerial transport better than bacteria due to their innate resistance to stressful physical conditions (UV radiation, desiccation, etc.). Survival of airborne cells might be more likely the result of an innate resistance (like fungal spores) than a resistance acquired while aerially transported. Available data support the hypothesis that planetary boundary layer microbial communities undergo a physical selection rather than genetic changes that allow a better survival and/or development once in the air. Moreover, microbial cells might face constantly changing conditions during aerial transport (changes in temperature, UV radiation, condensation/evaporation of water, etc.) that could prevent their adaptation. In the ocean, a faster evolution of microorganisms than their dispersal by ocean currents has been suggested in the Atlantic and Pacific oceans (oceanic surface current speed around 0.05 m/s, [123,124]). However, air currents could be 100 even 1000 times faster than surface oceanic currents. Inputs of new cells through aerosolization from Earth surfaces are significant and continuous in the planetary boundary layer.
4. Potential Impacts of Airborne Microbial Activity on Atmospheric Chemistry
If surviving microbial cells (both resistant and protected cells in particulate matter) remain active and even undergo a microbial adaptation to the physical and chemical conditions of the atmosphere while airborne, they might impact the atmospheric chemistry [18,19]. Planetary boundary layer microbial activity is expected to be quantitatively the most important within the atmosphere, as bioaerosols are mainly concentrated in this layer. Yet, atmospheric physical conditions such as UV radiation and desiccation might significantly affect the metabolic potential of the living microbial cells. UV radiation has been shown to be a critical factor restraining microbial activity of the oceanic surface bacterioplankton [125,126,127,128,129,130]. Irradiance affects bacterioplankton the most in spring and summer [129] and microbial activity was shown to be suppressed by up to 40% in the top five meters of the water column in near shore waters [126]. Airborne microbial activity might have a greater impact at night during which UV radiation does not reach the Earth’s surface and air relative humidity is typically higher when compared with daytime. Airborne microbial activity laboratory investigations have mainly been carried out on microorganisms isolated from cloud water and showed that microorganisms can degrade typical carboxylic compounds found in the air [18,20,36] (formate, acetate, formaldehyde, etc., [62]) as well as influence the oxidative capacity of clouds through the reduction of oxidants like H2O2 [21]. Using a liquid medium mimicking the composition of cloud water and a temperature of 5 °C (average temperature of low-altitude clouds), biological activity was shown to drive the oxidation of carbonaceous compounds during the night (90 to 99%), while contributing 2 to 37% of the reactivity during the day alongside radical reactions mediated by photochemistry [20]. rRNA-based studies identified the taxonomy of the potentially active microbial taxa in the dry troposphere and cloud water [62,131,132]. Epiphytic, parasitic and endosymbiont bacterial taxa (i.e., Sphingomonas, Methylobacterium, Acidiphilium, Pseudomonas, and Comamonas) have been suggested as the most active organisms due to their physiological properties (resistance to temperature and humidity shifts, high levels of UV radiation, etc.) compatible with their maintenance in the dry troposphere and clouds [38,131]. The same was observed for fungi with plant pathogens and saprophytic taxa (Pleosporales, Magnaporthales, Xylariales, Conioscyphales, etc.) potentially showing the highest activities [38,131]. Airborne microbial growth and reproduction have been suggested in cloud water [62,133]. Sattler et al. [133] suggested that bacterial division in cloud water might range from 3.6 to 19.5 days (production measurement at 0 °C), which was comparable to those of phytoplankton in the ocean, i.e., about a week [134]. Temperature in the planetary boundary layer might be higher than 0 °C and consequently, the microbial replication time might be less than 4 days. Residence time in the air might be a critical factor for planetary boundary layer microorganisms to divide, as microbial replication time might be on the same order as residence time. Most of the studies evaluating the metabolic potential of airborne microbial communities are based on cultivable microorganisms, and the conditions (physical and chemical) in which these experiments are carried out are far from those found in the atmosphere. Moreover, airborne microbial activity-related investigations have been mainly carried out on microorganisms isolated from cloud water where chemical species are in solution. Although a high diversity in functional genes has been revealed from planetary boundary layer microbial metagenomes [135,136], (Tignat-Perrier et al., in revision), the significance of planetary boundary layer microbial activity on atmospheric chemistry remains unknown.
5. Conclusions
The current development of culture-independent investigations in aeromicrobiology is producing valuable knowledge regarding the microbial ecology of airborne microbial communities. Yet heterogeneity in methodology (see [137,138,139,140] for recent methodology development and methodology comparison in aeromicrobiology) remains a major issue that needs to be considered when making comparisons between investigations. The different sampling strategies, especially the sample collection duration (a few hours, 24 h, one consecutive week, etc.), principle of collection (i.e., filtration, impaction, impingement in liquid), and size cut-offs (PM2.5, PM10, etc.) can lead to different interpretations. The principle of collection is associated to different DNA extraction methods, which both might result in a DNA recovery of different quality and yield that would subsequently affect sequencing outputs. Most available data strongly support the significant contribution of local sources (i.e., surrounding landscapes) and meteorology in the composition of planetary boundary layer microbial communities while still recent observations suggest randomization [141]. Physical selection of microorganisms is likely going on during aerosolization and aerial transport, and might contribute in selecting specific microorganisms from the underlying planetary bound ecosystems. Thus, planetary boundary layer microbial communities might not be the sum of the microbial communities of the different surrounding sources (soil, plants, etc.), but selected microorganisms that are more resistant to atmospheric conditions (desiccation, UV radiation, etc.), more aerosolized (due to membrane properties for example), and/or protected in atmospheric particulate matter. A better understanding of the physical selection (i.e., how atmospheric chemicals and physics influence microbial survival depending on microbial taxon and microbial distribution size) occurring during aerosolization and aerial transport is needed to be able to model the composition of planetary boundary layer microbial communities based on the surrounding landscapes and meteorological conditions. Although the activity of microorganisms of airborne origin has been shown on culture medium, airborne microbial activity should be evaluated in situ, i.e., in the field or under more controlled settings such as in atmospheric chambers. In situ activity, growth and microbial adaptation of airborne microorganisms might occur but to date no available data support them. Complex microbial communities (and not individual strains) should be investigated under controlled environmental conditions in atmospheric chambers that can mimic atmospheric conditions. These investigations might help detect if microbial communities are active, grow, and even undergo an adaption while airborne. Meta-omic investigations, especially metatranscriptomics and metaproteomics are currently very limited approaches (one study exists to date on cloud metatranscriptomics, see [37]) in aeromicrobiology due to the low biomass represented by airborne microorganisms and the sensitivity of sequencing technologies. Yet, these approaches might be useful for evaluating airborne microbial activity and should be further developed.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4433/11/12/1296/s1.
Author Contributions
A.D., C.L., T.M.V., and R.T.-P. had the idea for the article, R.T.-P. performed the literature search, data analysis, and wrote the first draft, all the authors critically revised the work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Agence Nationale de la Recherche (ANR-15-CE01-0002–03 INHALE); Région Auvergne-Rhône Alpes (ARC3 2016); CAMPUS France (program XU GUANGQI) and the French Polar Institute IPEV (program 1028 and 399).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Després, V.R.; Nowoisky, J.F.; Klose, M.; Conrad, R.; Andreae, M.O.; Pöschl, U. Characterization of primary biogenic aerosol particles in urban, rural, and high-alpine air by DNA sequence and restriction fragment analysis of ribosomal RNA genes. Biogeosciences 2007, 4, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Després, V.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.; Buryak, G.; Fröhlich-Nowoisky, J.; Elbert, W.; Andreae, M.; Pöschl, U.; et al. Primary biological aerosol particles in the atmosphere: A review. Tellus B Chem. Phys. Meteorol. 2012, 64, 15598. [Google Scholar] [CrossRef] [Green Version]
- Burrows, S.M.; Elbert, W.; Lawrence, M.G.; Pöschl, U. Bacteria in the global atmosphere—Part 1: Review and synthesis of literature data for different ecosystems. Atmos. Chem. Phys. 2009, 9, 9263–9280. [Google Scholar] [CrossRef] [Green Version]
- Burrows, S.M.; Butler, T.; Jöckel, P.; Tost, H.; Kerkweg, A.; Pöschl, U.; Lawrence, M.G. Bacteria in the global atmosphere—Part 2: Modeling of emissions and transport between different ecosystems. Atmos. Chem. Phys. 2009, 9, 9281–9297. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.J.; Griffin, D.W.; Jaffe, D.A. The high life: Transport of microbes in the atmosphere. Eos. Trans. AGU 2011, 92, 249–250. [Google Scholar] [CrossRef]
- Smith, D.J.; Ravichandar, J.D.; Jain, S.; Griffin, D.W.; Yu, H.; Tan, Q.; Thissen, J.; Lusby, T.; Nicoll, P.; Shedler, S.; et al. Airborne Bacteria in Earth’s Lower Stratosphere Resemble Taxa Detected in the Troposphere: Results From a New NASA Aircraft Bioaerosol Collector (ABC). Front. Microbiol 2018, 9, 1752. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.W. Terrestrial Microorganisms at an Altitude of 20,000 m in Earth’s Atmosphere. Aerobiologia 2004, 20, 135–140. [Google Scholar] [CrossRef]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.W. Atmospheric Movement of Microorganisms in Clouds of Desert Dust and Implications for Human Health. Clin. Microbiol. Rev. 2007, 20, 459–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wainwright, M.; Wickramasinghe, N.C.; Narlikar, J.V.; Rajaratnam, P. Microorganisms cultured from stratospheric air samples obtained at 41 km. FEMS Microbiol. Lett. 2003, 218, 161–165. [Google Scholar] [CrossRef]
- Imshenetsky, A.A.; Lysenko, S.V.; Kazakov, G.A. Upper boundary of the biosphere. Appl. Environ. Microbiol. 1978, 35, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.W.; Gonzalez-Martin, C.; Hoose, C.; Smith, D.J. Global-Scale Atmospheric Dispersion of Microorganisms. In Microbiology of Aerosols; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 155–194. ISBN 978-1-119-13231-8. [Google Scholar]
- Smith, D.J. Aeroplankton and the Need for a Global Monitoring Network. BioScience 2013, 63, 515–516. [Google Scholar] [CrossRef] [Green Version]
- Delort, A.-M.; Vaïtilingom, M.; Amato, P.; Sancelme, M.; Parazols, M.; Mailhot, G.; Laj, P.; Deguillaume, L. A short overview of the microbial population in clouds: Potential roles in atmospheric chemistry and nucleation processes. Atmos. Res. 2010, 98, 249–260. [Google Scholar] [CrossRef]
- Failor, K.C.; Schmale, D.G.; Vinatzer, B.A.; Monteil, C.L. Ice nucleation active bacteria in precipitation are genetically diverse and nucleate ice by employing different mechanisms. ISME J. 2017, 11, 2740–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariya, P.; Sun, J.; Eltouny, N.; Hudson, E.; Hayes, C.; Kos, G. Physical and chemical characterization of bioaerosols--Implications for nucleation processes. Int. Rev. Phys. Chem. 2009, 28, 1–32. [Google Scholar] [CrossRef]
- Haga, D.I.; Burrows, S.M.; Iannone, R.; Wheeler, M.J.; Mason, R.H.; Chen, J.; Polishchuk, E.A.; Pöschl, U.; Bertram, A.K. Ice nucleation by fungal spores from the classes Agaricomycetes, Ustilaginomycetes, and Eurotiomycetes, and the effect on the atmospheric transport of these spores. Atmos. Chem. Phys. 2014, 14, 8611–8630. [Google Scholar] [CrossRef] [Green Version]
- Ariya, P.A.; Nepotchatykh, O.; Ignatova, O.; Amyot, M. Microbiological degradation of atmospheric organic compounds. Geophys. Res. Lett. 2002, 29, 34–1–34–4. [Google Scholar] [CrossRef] [Green Version]
- Ariya, P.A.; Amyot, M. New Directions: The role of bioaerosols in atmospheric chemistry and physics. Atmos. Environ. 2004, 38, 1231–1232. [Google Scholar] [CrossRef]
- Vaïtilingom, M.; Amato, P.; Sancelme, M.; Laj, P.; Leriche, M.; Delort, A.-M. Contribution of Microbial Activity to Carbon Chemistry in Clouds. Appl. Environ. Microbiol. 2010, 76, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Vaïtilingom, M.; Deguillaume, L.; Vinatier, V.; Sancelme, M.; Amato, P.; Chaumerliac, N.; Delort, A.-M. Potential impact of microbial activity on the oxidant capacity and organic carbon budget in clouds. Proc. Natl. Acad. Sci. USA 2013, 110, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Aylor, D.E. Spread of Plant Disease on a Continental Scale: Role of Aerial Dispersal of Pathogens. Ecology 2003, 84, 1989–1997. [Google Scholar] [CrossRef]
- Lee, B. Life Comes from the Air: A Short Review on Bioaerosol Control. Aerosol Air Qual. Res. 2011, 11. [Google Scholar] [CrossRef] [Green Version]
- Srikanth, P.; Sudharsanam, S.; Steinberg, R. Bio-aerosols in indoor environment: Composition, health effects and analysis. Indian J. Med. Microbiol. 2008, 26, 302–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, V.; Gandolfi, I.; Ambrosini, R.; Bestetti, G.; Innocente, E.; Rampazzo, G.; Franzetti, A. Temporal variability and effect of environmental variables on airborne bacterial communities in an urban area of Northern Italy. Appl. Microbiol. Biotechnol. 2013, 97, 6561–6570. [Google Scholar] [CrossRef] [Green Version]
- Zhen, Q.; Deng, Y.; Wang, Y.; Wang, X.; Zhang, H.; Sun, X.; Ouyang, Z. Meteorological factors had more impact on airborne bacterial communities than air pollutants. Sci. Total Environ. 2017, 601–602, 703–712. [Google Scholar] [CrossRef]
- Genitsaris, S.; Stefanidou, N.; Katsiapi, M.; Kormas, K.A.; Sommer, U.; Moustaka-Gouni, M. Variability of airborne bacteria in an urban Mediterranean area (Thessaloniki, Greece). Atmos. Environ. 2017, 157, 101–110. [Google Scholar] [CrossRef]
- Gandolfi, I.; Bertolini, V.; Bestetti, G.; Ambrosini, R.; Innocente, E.; Rampazzo, G.; Papacchini, M.; Franzetti, A. Spatio-temporal variability of airborne bacterial communities and their correlation with particulate matter chemical composition across two urban areas. Appl. Microbiol. Biotechnol. 2015, 99, 4867–4877. [Google Scholar] [CrossRef]
- Cho, B.C.; Hwang, C.Y. Prokaryotic abundance and 16S rRNA gene sequences detected in marine aerosols on the East Sea (Korea). FEMS Microbiol. Ecol. 2011, 76, 327–341. [Google Scholar] [CrossRef]
- Park, J.; Li, P.-F.; Ichijo, T.; Nasu, M.; Yamaguchi, N. Effects of Asian dust events on atmospheric bacterial communities at different distances downwind of the source region. J. Environ. Sci. 2018, 72, 133–139. [Google Scholar] [CrossRef]
- Tanaka, D.; Sato, K.; Goto, M.; Fujiyoshi, S.; Maruyama, F.; Takato, S.; Shimada, T.; Sakatoku, A.; Aoki, K.; Nakamura, S. Airborne Microbial Communities at High-Altitude and Suburban Sites in Toyama, Japan Suggest a New Perspective for Bioprospecting. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Zweifel, U.L.; Hagström, Å.; Holmfeldt, K.; Thyrhaug, R.; Geels, C.; Frohn, L.M.; Skjøth, C.A.; Karlson, U.G. High bacterial 16S rRNA gene diversity above the atmospheric boundary layer. Aerobiologia 2012, 28, 481–498. [Google Scholar] [CrossRef]
- DeLeon-Rodriguez, N. Microbiome of the Upper Troposphere: Species Composition and Prevalence, Effects of Tropical Storms, and Atmospheric Implications. Available online: http://www.pnas.org/content/110/7/2575.full (accessed on 25 July 2017).
- Els, N.; Larose, C.; Baumann-Stanzer, K.; Tignat-Perrier, R.; Keuschnig, C.; Vogel, T.M.; Sattler, B. Microbial composition in seasonal time series of free tropospheric air and precipitation reveals community separation. Aerobiologia 2019. [Google Scholar] [CrossRef] [Green Version]
- Amato, P.; Ménager, M.; Sancelme, M.; Laj, P.; Mailhot, G.; Delort, A.-M. Microbial population in cloud water at the Puy de Dôme: Implications for the chemistry of clouds. Atmos. Environ. 2005, 39, 4143–4153. [Google Scholar] [CrossRef] [Green Version]
- Amato, P.; Besaury, L.; Joly, M.; Penaud, B.; Deguillaume, L.; Delort, A.-M. Metatranscriptomic exploration of microbial functioning in clouds. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Amato, P.; Joly, M.; Besaury, L.; Oudart, A.; Taib, N.; Moné, A.I.; Deguillaume, L.; Delort, A.-M.; Debroas, D. Active microorganisms thrive among extremely diverse communities in cloud water. PLoS ONE 2017, 12, e0182869. [Google Scholar] [CrossRef] [Green Version]
- Renard, P.; Canet, I.; Sancelme, M.; Wirgot, N.; Deguillaume, L.; Delort, A.-M. Screening of cloud microorganisms isolated at the Puy de Dôme (France) station for the production of biosurfactants. Atmos. Chem. Phys. 2016, 16, 12347–12358. [Google Scholar] [CrossRef] [Green Version]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Bowers, R.M.; McCubbin, I.B.; Hallar, A.G.; Fierer, N. Seasonal variability in airborne bacterial communities at a high-elevation site. Atmos. Environ. 2012, 50, 41–49. [Google Scholar] [CrossRef]
- Bowers, R.M.; Clements, N.; Emerson, J.B.; Wiedinmyer, C.; Hannigan, M.P.; Fierer, N. Seasonal variability in bacterial and fungal diversity of the near-surface atmosphere. Environ. Sci. Technol. 2013, 47, 12097–12106. [Google Scholar] [CrossRef] [PubMed]
- Mayol, E.; Arrieta, J.M.; Jiménez, M.A.; Martínez-Asensio, A.; Garcias-Bonet, N.; Dachs, J.; González-Gaya, B.; Royer, S.-J.; Benítez-Barrios, V.M.; Fraile-Nuez, E.; et al. Long-range transport of airborne microbes over the global tropical and subtropical ocean. Nat. Commun. 2017, 8, 201. [Google Scholar] [CrossRef]
- Fulton, J.D. Microorganisms of the Upper Atmosphere: III. Relationship between Altitude and Micropopulation. Appl. Environ. Microbiol. 1966, 14, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Els, N.; Baumann-Stanzer, K.; Larose, C.; Vogel, T.M.; Sattler, B. Beyond the planetary boundary layer: Bacterial and fungal vertical biogeography at Mount Sonnblick, Austria. Geo Geogr. Environ. 2019, 6, e00069. [Google Scholar] [CrossRef]
- Griffin, D.W.; Kellogg, C.A.; Garrison, V.H.; Lisle, J.T.; Borden, T.C.; Shinn, E.A. Atmospheric microbiology in the northern Caribbean during African dust events. Aerobiologia 2003, 19, 143–157. [Google Scholar] [CrossRef]
- Li, Y.; Lu, R.; Li, W.; Xie, Z.; Song, Y. Concentrations and size distributions of viable bioaerosols under various weather conditions in a typical semi-arid city of Northwest China. J. Aerosol Sci. 2017, 106, 83–92. [Google Scholar] [CrossRef]
- Weil, T.; De Filippo, C.; Albanese, D.; Donati, C.; Pindo, M.; Pavarini, L.; Carotenuto, F.; Pasqui, M.; Poto, L.; Gabrieli, J.; et al. Legal immigrants: Invasion of alien microbial communities during winter occurring desert dust storms. Microbiome 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, N.; Baba, T.; Ichijo, T.; Himezawa, Y.; Enoki, K.; Saraya, M.; Li, P.-F.; Nasu, M. Abundance and Community Structure of Bacteria on Asian Dust Particles Collected in Beijing, China, during the Asian Dust Season. Biol. Pharm. Bull. 2016, 39, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Qi, J.; Shao, C.; Zhong, X.; Gao, D.; Cao, W.; Gao, J.; Bai, R.; Long, G.; Chu, C. Concentration and size distribution of total airborne microbes in hazy and foggy weather. Sci. Total Environ. 2016, 541, 1011–1018. [Google Scholar] [CrossRef]
- Clauss, M. Particle size distribution of airborne micro-organisms in the environment-A review. Landbauforsch. Volkenrode 2015, 65, 77–100. [Google Scholar] [CrossRef]
- Amato, P.; Joly, M.; Schaupp, C.; Attard, E.; Möhler, O.; Morris, C.E.; Brunet, Y.; Delort, A.-M. Survival and ice nucleation activity of bacteria as aerosols in a cloud simulation chamber. Atmos. Chem. Phys. 2015, 15, 6455–6465. [Google Scholar] [CrossRef]
- Mayol, E.; Jiménez, M.A.; Herndl, G.J.; Duarte, C.M.; Arrieta, J.M. Resolving the abundance and air-sea fluxes of airborne microorganisms in the North Atlantic Ocean. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Aller, J.Y.; Kuznetsova, M.R.; Jahns, C.J.; Kemp, P.F. The sea surface microlayer as a source of viral and bacterial enrichment in marine aerosols. J. Aerosol Sci. 2005, 36, 801–812. [Google Scholar] [CrossRef]
- Blanchard, D.C.; Syzdek, L.D. Water-to-Air Transfer and Enrichment of Bacteria in Drops from Bursting Bubbles. Appl. Environ. Microbiol. 1982, 43, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Seinfeld, J.H.; Pandis, S.N. Environmental Chemistry—Chemistry—Subjects—Wiley. In Atmospheric Chemistry and Physics: From Air Pollution to Climate Change, 3rd ed.; Wiley-Interscience: Hoboken, NJ, USA, 1998. [Google Scholar]
- Jaenicke, R. Atmospheric aerosols and global climate. J. Aerosol Sci. 1980, 11, 577–588. [Google Scholar] [CrossRef]
- Tignat-Perrier, R.; Dommergue, A.; Thollot, A.; Keuschnig, C.; Magand, O.; Vogel, T.M.; Larose, C. Global airborne microbial communities controlled by surrounding landscapes and wind conditions. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Michaud, J.M.; Thompson, L.R.; Kaul, D.; Espinoza, J.L.; Richter, R.A.; Xu, Z.Z.; Lee, C.; Pham, K.M.; Beall, C.M.; Malfatti, F.; et al. Taxon-specific aerosolization of bacteria and viruses in an experimental ocean-atmosphere mesocosm. Nat. Commun. 2018, 9, 2017. [Google Scholar] [CrossRef]
- Fahlgren, C.; Gómez-Consarnau, L.; Zábori, J.; Lindh, M.V.; Krejci, R.; Mårtensson, E.M.; Nilsson, D.; Pinhassi, J. Seawater mesocosm experiments in the Arctic uncover differential transfer of marine bacteria to aerosols. Environ. Microbiol. Rep. 2015, 7, 460–470. [Google Scholar] [CrossRef]
- Amato, P.; Demeer, F.; Melaouhi, A.; Fontanella, S.; Martin-Biesse, A.-S.; Sancelme, M.; Laj, P.; Delort, A.-M. A fate for organic acids, formaldehyde and methanol in cloud water: Their biotransformation by micro-organisms. Atmos. Chem. Phys. 2007, 7, 4159–4169. [Google Scholar] [CrossRef] [Green Version]
- Alghamdi, M.A.; Shamy, M.; Redal, M.A.; Khoder, M.; Awad, A.H.; Elserougy, S. Microorganisms associated particulate matter: A preliminary study. Sci. Total Environ. 2014, 479–480, 109–116. [Google Scholar] [CrossRef]
- Womack, A.M.; Artaxo, P.E.; Ishida, F.Y.; Mueller, R.C.; Saleska, S.R.; Wiedemann, K.T.; Bohannan, B.J.M.; Green, J.L. Characterization of active and total fungal communities in the atmosphere over the Amazon rainforest. Biogeosciences 2015, 12, 6337–6349. [Google Scholar] [CrossRef] [Green Version]
- Sesartic, A.; Lohmann, U.; Storelvmo, T. Bacteria in the ECHAM5-HAM global climate model. Atmos. Chem. Phys. 2012, 12, 8645–8661. [Google Scholar] [CrossRef] [Green Version]
- Elbert, W.; Taylor, P.E.; Andreae, M.O.; Pöschl, U. Contribution of fungi to primary biogenic aerosols in the atmosphere: Wet and dry discharged spores, carbohydrates, and inorganic ions. Atmos. Chem. Phys. 2007, 7, 4569–4588. [Google Scholar] [CrossRef] [Green Version]
- Crandall, S.G.; Gilbert, G.S. Meteorological factors associated with abundance of airborne fungal spores over natural vegetation. Atmos. Environ. 2017, 162, 87–99. [Google Scholar] [CrossRef]
- Górny, R.; Lawniczek-Walczyk, A. Effect of two aerosolization methods on the release of fungal propagules from a contaminated agar surface. Ann. Agric. Environ. Med. AAEM 2012, 19, 279–284. [Google Scholar]
- Joung, Y.S.; Ge, Z.; Buie, C.R. Bioaerosol generation by raindrops on soil. Nat. Commun. 2017, 8, 14668. [Google Scholar] [CrossRef]
- Zhai, Y.; Li, X.; Wang, T.; Wang, B.; Li, C.; Zeng, G. A review on airborne microorganisms in particulate matters: Composition, characteristics and influence factors. Environ. Int. 2018, 113, 74–90. [Google Scholar] [CrossRef]
- Bowers, R.M.; McLetchie, S.; Knight, R.; Fierer, N. Spatial variability in airborne bacterial communities across land-use types and their relationship to the bacterial communities of potential source environments. ISME J. 2011, 5, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Innocente, E.; Squizzato, S.; Visin, F.; Facca, C.; Rampazzo, G.; Bertolini, V.; Gandolfi, I.; Franzetti, A.; Ambrosini, R.; Bestetti, G. Influence of seasonality, air mass origin and particulate matter chemical composition on airborne bacterial community structure in the Po Valley, Italy. Sci. Total Environ. 2017, 677–687. [Google Scholar] [CrossRef]
- Uetake, J.; Tobo, Y.; Uji, Y.; Hill, T.C.J.; DeMott, P.J.; Kreidenweis, S.; Misumi, R. Seasonal changes of airborne bacterial communities over Tokyo and influence of local meteorology. arXiv 2019, arXiv:Bio/542001. [Google Scholar] [CrossRef] [Green Version]
- Tignat-Perrier, R.; Dommergue, A.; Thollot, A.; Magand, O.; Amato, P.; Joly, M.; Sellegri, K.; Vogel, T.M.; Larose, C. Seasonal shift in airborne microbial communities. Sci. Total Environ. 2020, 716, 137129. [Google Scholar] [CrossRef] [PubMed]
- Maki, T.; Hara, K.; Kobayashi, F.; Kurosaki, Y.; Kakikawa, M.; Matsuki, A.; Chen, B.; Shi, G.; Hasegawa, H.; Iwasaka, Y. Vertical distribution of airborne bacterial communities in an Asian-dust downwind area, Noto Peninsula. Atmos. Environ. 2015, 119, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Gat, D.; Mazar, Y.; Cytryn, E.; Rudich, Y. Origin-Dependent Variations in the Atmospheric Microbiome Community in Eastern Mediterranean Dust Storms. Environ. Sci. Technol. 2017, 51, 6709–6718. [Google Scholar] [CrossRef] [PubMed]
- Maki, T.; Hara, K.; Iwata, A.; Lee, K.C.; Kawai, K.; Kai, K.; Kobayashi, F.; Pointing, S.B.; Archer, S.; Hasegawa, H.; et al. Variations in airborne bacterial communities at high altitudes over the Noto Peninsula (Japan) in response to Asian dust events. Atmos. Chem. Phys. 2017, 17, 11877–11897. [Google Scholar] [CrossRef] [Green Version]
- Reche, I.; Ortega-Retuerta, E.; Romera, O.; Villena, E.P.; Baquero, R.M.; Casamayor, E.O. Effect of Saharan dust inputs on bacterial activity and community composition in Mediterranean lakes and reservoirs. Limnol. Oceanogr. 2009, 54, 869–879. [Google Scholar] [CrossRef] [Green Version]
- Prospero, J.M.; Blades, E.; Mathison, G.; Naidu, R. Interhemispheric transport of viable fungi and bacteria from Africa to the Caribbean with soil dust. Aerobiologia 2005, 21, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Creamean, J.M.; Suski, K.J.; Rosenfeld, D.; Cazorla, A.; DeMott, P.J.; Sullivan, R.C.; White, A.B.; Ralph, F.M.; Minnis, P.; Comstock, J.M.; et al. Dust and Biological Aerosols from the Sahara and Asia Influence Precipitation in the Western U.S. Science 2013, 339, 1572–1578. [Google Scholar] [CrossRef] [Green Version]
- Itani, G.N.; Smith, C.A. Dust Rains Deliver Diverse Assemblages of Microorganisms to the Eastern Mediterranean. Sci. Rep. 2016, 6, 22657. [Google Scholar] [CrossRef]
- Maki, T.; Susuki, S.; Kobayashi, F.; Kakikawa, M.; Tobo, Y.; Yamada, M.; Higashi, T.; Matsuki, A.; Hong, C.; Hasegawa, H.; et al. Phylogenetic analysis of atmospheric halotolerant bacterial communities at high altitude in an Asian dust (KOSA) arrival region, Suzu City. Sci. Total Environ. 2010, 408, 4556–4562. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.W.; Garrison, V.H.; Herman, J.R.; Shinn, E.A. African desert dust in the Caribbean atmosphere: Microbiology and public health. Aerobiologia 2001, 17, 203–213. [Google Scholar] [CrossRef]
- Kellogg, C.A.; Griffin, D.W.; Garrison, V.H.; Peak, K.K.; Royall, N.; Smith, R.R.; Shinn, E.A. Characterization of Aerosolized Bacteria and Fungi From Desert Dust Events in Mali, West Africa. Aerobiologia 2004, 20, 99–110. [Google Scholar] [CrossRef]
- Maki, T.; Lee, K.C.; Kawai, K.; Onishi, K.; Hong, C.S.; Kurosaki, Y.; Shinoda, M.; Kai, K.; Iwasaka, Y.; Archer, S.D.J.; et al. Aeolian Dispersal of Bacteria Associated With Desert Dust and Anthropogenic Particles Over Continental and Oceanic Surfaces. J. Geophys. Res. Atmos. 2019, 124, 5579–5588. [Google Scholar] [CrossRef]
- Katra, I.; Arotsker, L.; Krasnov, H.; Zaritsky, A.; Kushmaro, A.; Ben-Dov, E. Richness and Diversity in Dust Stormborne Biomes at the Southeast Mediterranean. Sci. Rep. 2014, 4, 5265. [Google Scholar] [CrossRef] [PubMed]
- Delmont, T.O.; Malandain, C.; Prestat, E.; Larose, C.; Monier, J.-M.; Simonet, P.; Vogel, T.M. Metagenomic mining for microbiologists. ISME J. 2011, 5, 1837–1843. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Wang, F.; Guo, L.; Chen, Z.; Sievert, S.M.; Meng, J.; Huang, G.; Li, Y.; Yan, Q.; Wu, S.; et al. Comparative metagenomics of microbial communities inhabiting deep-sea hydrothermal vent chimneys with contrasting chemistries. ISME J. 2011, 5, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, L.; Zhang, Y.; Liu, H.; Jing, H. Comparative metagenomics study reveals pollution induced changes of microbial genes in mangrove sediments. Sci. Rep. 2019, 9, 5739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tringe, S.G.; von Mering, C.; Kobayashi, A.; Salamov, A.A.; Chen, K.; Chang, H.W.; Podar, M.; Short, J.M.; Mathur, E.J.; Detter, J.C.; et al. Comparative metagenomics of microbial communities. Science 2005, 308, 554–557. [Google Scholar] [CrossRef] [Green Version]
- Moan, J.; Peak, M.J. Effects of UV radiation of cells. J. Photochem. Photobiol. B Biol. 1989, 4, 21–34. [Google Scholar] [CrossRef]
- Santos, A.L.; Oliveira, V.; Baptista, I.; Henriques, I.; Gomes, N.C.M.; Almeida, A.; Correia, A.; Cunha, Â. Wavelength dependence of biological damage induced by UV radiation on bacteria. Arch. Microbiol. 2013, 195, 63–74. [Google Scholar] [CrossRef]
- DasSarma, P.; DasSarma, S. Survival of microbes in Earth’s stratosphere. Curr. Opin. Microbiol. 2018, 43, 24–30. [Google Scholar] [CrossRef]
- Smith, D.J.; Griffin, D.W.; McPeters, R.D.; Ward, P.D.; Schuerger, A.C. Microbial survival in the stratosphere and implications for global dispersal. Aerobiologia 2011, 27, 319–332. [Google Scholar] [CrossRef]
- Yang, Y.; Yokobori, S.; Yamagishi, A. UV-resistant bacteria isolated from upper troposphere and lower stratosphere. Biol. Sci. Space 2008, 22, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Bryan, N.C.; Christner, B.C.; Guzik, T.G.; Granger, D.J.; Stewart, M.F. Abundance and survival of microbial aerosols in the troposphere and stratosphere. ISME J. 2019, 13, 2789–2799. [Google Scholar] [CrossRef] [PubMed]
- Rangel, D.E.N.; Braga, G.U.L.; Fernandes, É.K.K.; Keyser, C.A.; Hallsworth, J.E.; Roberts, D.W. Stress tolerance and virulence of insect-pathogenic fungi are determined by environmental conditions during conidial formation. Curr. Genet. 2015, 61, 383–404. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Anderson, A.J.; Roberts, D.W. Evaluating physical and nutritional stress during mycelial growth as inducers of tolerance to heat and UV-B radiation in Metarhizium anisopliae conidia. Mycol. Res. 2008, 112, 1362–1372. [Google Scholar] [CrossRef]
- Hagiwara, D.; Sakai, K.; Suzuki, S.; Umemura, M.; Nogawa, T.; Kato, N.; Osada, H.; Watanabe, A.; Kawamoto, S.; Gonoi, T.; et al. Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Vorob’eva, L.I. Stressors, Stress Reactions, and Survival of Bacteria: A Review. Appl. Biochem. Microbiol. 2004, 40, 217–224. [Google Scholar] [CrossRef]
- D’Amico, S.; Collins, T.; Marx, J.-C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Nedwell, D.B. Effect of low temperature on microbial growth: Lowered affinity for substrates limits growth at low temperature. FEMS Microbiol. Ecol. 1999, 30, 101–111. [Google Scholar] [CrossRef]
- Berry, E.D.; Foegeding, P.M. Cold Temperature Adaptation and Growth of Microorganisms. J. Food Prot. 1997, 60, 1583–1594. [Google Scholar] [CrossRef]
- Joly, M.; Amato, P.; Sancelme, M.; Vinatier, V.; Abrantes, M.; Deguillaume, L.; Delort, A.-M. Survival of microbial isolates from clouds toward simulated atmospheric stress factors. Atmos. Environ. 2015, 117, 92–98. [Google Scholar] [CrossRef]
- Ehrlich, R.; Miller, S.; Walker, R.L. Effects of Atmospheric Humidity and Temperature on the Survival of Airborne Flavobacterium. Appl. Environ. Microbiol. 1970, 20, 884–887. [Google Scholar] [CrossRef] [Green Version]
- Wright, D.N.; Bailey, G.D.; Hatch, M.T. Survival of airborne Mycoplasma as affected by relative humidity. J. Bacteriol. 1968, 95, 251–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattimore, V.; Battista, J.R. Radioresistance of Deinococcus radiodurans: Functions necessary to survive ionizing radiation are also necessary to survive prolonged desiccation. J. Bacteriol. 1996, 178, 633–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dose, K.; Bieger-Dose, A.; Labusch, M.; Gill, M. Survival in extreme dryness and DNA-single-strand breaks. Adv. Space Res. 1992, 12, 221–229. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Li, S.W.; Gaidamakova, E.K.; Matrosova, V.Y.; Zhai, M.; Sulloway, H.M.; Scholten, J.C.; Brown, M.G.; Balkwill, D.L.; Daly, M.J. Protein oxidation: Key to bacterial desiccation resistance? ISME J. 2008, 2, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredrickson, J.K.; Zachara, J.M.; Balkwill, D.L.; Kennedy, D.; Li, S.W.; Kostandarithes, H.M.; Daly, M.J.; Romine, M.F.; Brockman, F.J. Geomicrobiology of High-Level Nuclear Waste-Contaminated Vadose Sediments at the Hanford Site, Washington State. Appl. Environ. Microbiol. 2004, 70, 4230–4241. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.W.; Maxcy, R.B. Isolation of radiation-resistant bacteria without exposure to irradiation. Appl. Environ. Microbiol. 1979, 38, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Rainey, F.A.; Ray, K.; Ferreira, M.; Gatz, B.Z.; Nobre, M.F.; Bagaley, D.; Rash, B.A.; Park, M.-J.; Earl, A.M.; Shank, N.C.; et al. Extensive Diversity of Ionizing-Radiation-Resistant Bacteria Recovered from Sonoran Desert Soil and Description of Nine New Species of the Genus Deinococcus Obtained from a Single Soil Sample. Appl. Environ. Microbiol. 2005, 71, 5225–5235. [Google Scholar] [CrossRef] [Green Version]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.M. Bacterial responses to osmotic challenges. J. Gen. Physiol. 2015, 145, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Alsved, M.; Holm, S.; Christiansen, S.; Smidt, M.; Ling, M.; Boesen, T.; Finster, K.; Bilde, M.; Löndahl, J.; Šantl-Temkiv, T. Effect of Aerosolization and Drying on the Viability of Pseudomonas syringae Cells. Front. Microbiol. 2018, 9, 3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deguillaume, L.; Charbouillot, T.; Joly, M.; Vaïtilingom, M.; Parazols, M.; Marinoni, A.; Amato, P.; Delort, A.-M.; Vinatier, V.; Flossmann, A.; et al. Classification of clouds sampled at the puy de Dôme (France) based on 10 yr of monitoring of their physicochemical properties. Atmos. Chem. Phys. 2014, 14, 1485–1506. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.J.; Webber, D.; Hopkins, R.; Frost, A.; Laws, T.; Jayasekera, P.N.; Atkins, T. The Cell Membrane as a Major Site of Damage during Aerosolization of Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 920–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, H.; Han, T.; Fennell, D.E.; Mainelis, G. Release of Free DNA by Membrane-Impaired Bacterial Aerosols Due to Aerosolization and Air Sampling. Appl. Environ. Microbiol. 2013, 79, 7780–7789. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.W.; Chan, W.L.; Lai, K.M. Importance of stress-response genes to the survival of airborne Escherichia coli under different levels of relative humidity. AMB Express 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.W.; Ip, M.; Chao, C.Y.H.; Tang, J.W.; Lai, K.P.; Fu, S.C.; Leung, W.T.; Lai, K.M. Differential gene expression in Escherichia coli during aerosolization from liquid suspension. Appl. Microbiol. Biotechnol. 2018, 102, 6257–6267. [Google Scholar] [CrossRef]
- Mainelis, G.; Górny, R.L.; Reponen, T.; Trunov, M.; Grinshpun, S.A.; Baron, P.; Yadav, J.; Willeke, K. Effect of electrical charges and fields on injury and viability of airborne bacteria. Biotechnol. Bioeng. 2002, 79, 229–241. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; Samson, R.A. Food Mycology: A Multifaceted Approach to Fungi and Food; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-1-4200-2098-4. [Google Scholar]
- Martiny, A.C.; Tai, A.P.K.; Veneziano, D.; Primeau, F.; Chisholm, S.W. Taxonomic resolution, ecotypes and the biogeography of Prochlorococcus. Environ. Microbiol. 2009, 11, 823–832. [Google Scholar] [CrossRef]
- Hellweger, F.L.; van Sebille, E.; Fredrick, N.D. Biogeographic patterns in ocean microbes emerge in a neutral agent-based model. Science 2014, 345, 1346–1349. [Google Scholar] [CrossRef]
- Hernández, K.L.; Quiñones, R.A.; Daneri, G.; Farias, M.E.; Helbling, E.W. Solar UV radiation modulates daily production and DNA damage of marine bacterioplankton from a productive upwelling zone (36° S), Chile. J. Exp. Mar. Biol. Ecol. 2007, 343, 82–95. [Google Scholar] [CrossRef]
- Herndl, G.J.; Müller-Niklas, G.; Frick, J. Major role of ultraviolet-B in controlling bacterioplankton growth in the surface layer of the ocean. Nature 1993, 361, 717–719. [Google Scholar] [CrossRef]
- Winter, C.; Moeseneder, M.M.; Herndl, G.J. Impact of UV Radiation on Bacterioplankton Community Composition. Appl. Environ. Microbiol. 2001, 67, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Sáez, L.; Gasol, J.M.; Lefort, T.; Hofer, J.; Sommaruga, R. Effect of natural sunlight on bacterial activity and differential sensitivity of natural bacterioplankton groups in northwestern Mediterranean coastal waters. Appl. Environ. Microbiol. 2006, 72, 5806–5813. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-González, C.; Lefort, T.; Galí, M.; Montserrat Sala, M.; Sommaruga, R.; Simó, R.; Gasol, J.M. Seasonal patterns in the sunlight sensitivity of bacterioplankton from Mediterranean surface coastal waters. FEMS Microbiol. Ecol. 2012, 79, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Gonzalez, C.; Simó, R.; Sommaruga, R.; Gasol, J.M. Away from darkness: A review on the effects of solar radiation on heterotrophic bacterioplankton activity. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.M.; Bohannan, B.J.M.; Jaffe, D.A.; Levin, D.A.; Green, J.L. Molecular Evidence for Metabolically Active Bacteria in the Atmosphere. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Womack, A.M. UV-resistant bacteria isolated from upper troposphere and lower stratosphere (PDF Download Available). ResearchGate 2010. [Google Scholar] [CrossRef] [Green Version]
- Sattler, B.; Puxbaum, H.; Psenner, R. Bacterial growth in supercooled cloud droplets. Geophys. Res. Lett. 2001, 28, 239–242. [Google Scholar] [CrossRef]
- Falkowski, P.; Barber, R.; Smetacek, V. Biogeochemical Controls and Feedbacks on Ocean Primary Production. Science 1998, 281, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Yooseph, S.; Andrews-Pfannkoch, C.; Tenney, A.; McQuaid, J.; Williamson, S.; Thiagarajan, M.; Brami, D.; Zeigler-Allen, L.; Hoffman, J.; Goll, J.B.; et al. A Metagenomic Framework for the Study of Airborne Microbial Communities. PLoS ONE 2013, 8, e81862. [Google Scholar] [CrossRef] [PubMed]
- Aalismail, N.A.; Ngugi, D.K.; Díaz-Rúa, R.; Alam, I.; Cusack, M.; Duarte, C.M. Functional metagenomic analysis of dust-associated microbiomes above the Red Sea. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bøifot, K.O.; Gohli, J.; Moen, L.V.; Dybwad, M. Performance evaluation of a new custom, multi-component DNA isolation method optimized for use in shotgun metagenomic sequencing-based aerosol microbiome research. Environ. Microbiome 2020, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Dommergue, A.; Amato, P.; Tignat-Perrier, R.; Magand, O.; Thollot, A.; Joly, M.; Bouvier, L.; Sellegri, K.; Vogel, T.; Sonke, J.E.; et al. Methods to investigate the global atmospheric microbiome. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Bilodeau, G.J.; Duchaine, C. Bioaerosol Sampler Choice Should Consider Efficiency and Ability of Samplers To Cover Microbial Diversity. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, E.; Johansson, A.-M.; Ahlinder, J.; Lundkvist, M.J.; Singh, N.J.; Brodin, T.; Forsman, M.; Stenberg, P. Airborne microbial biodiversity and seasonality in Northern and Southern Sweden. PeerJ 2020, 8, e8424. [Google Scholar] [CrossRef] [Green Version]
- Tipton, L.; Zahn, G.; Datlof, E.; Kivlin, S.N.; Sheridan, P.; Amend, A.S.; Hynson, N.A. Fungal aerobiota are not affected by time nor environment over a 13-y time series at the Mauna Loa Observatory. Proc. Natl. Acad. Sci. USA 2019, 116, 25728–25733. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The different biological niches investigated in aeromicrobiology. These different niches might exchange microorganisms and represent different physico-chemical conditions for airborne microorganisms.
Figure 1.
The different biological niches investigated in aeromicrobiology. These different niches might exchange microorganisms and represent different physico-chemical conditions for airborne microorganisms.

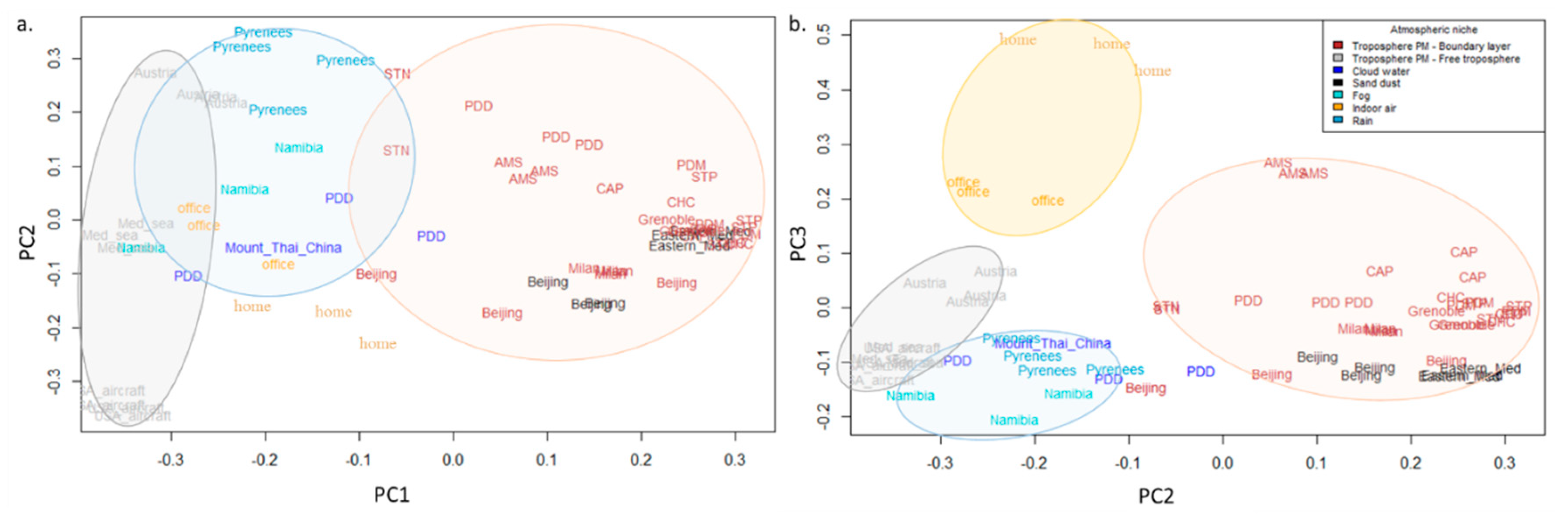
Figure 2.
Distribution of the samples coming from different phases of the atmosphere (cloud water, rain, fog, planetary boundary layer, and free troposphere particulate matter) based on the bacterial community structure. Public 16S rRNA gene sequencing based datasets were downloaded on public databases (Table S1 in Supplementary Data). Reads were filtered based on quality using FASTX-Toolkit then PANDAseq [40] was used to assemble the read 1 and read 2 using the RDP algorithm (Ramer–Douglas–Peucker algorithm). The resulting sequences were annotated at the genus level by RDP Classifier [41] using the RDP 16srrna database and an assignment confidence cutoff of 0.6. The PCo (Principal coordinates) analysis (components 1 and 2 in (a), components 2 and 3 in (b)) of the Bray–Curtis dissimilarity matrix is based on the relative abundances of the different bacterial genera.
Figure 2.
Distribution of the samples coming from different phases of the atmosphere (cloud water, rain, fog, planetary boundary layer, and free troposphere particulate matter) based on the bacterial community structure. Public 16S rRNA gene sequencing based datasets were downloaded on public databases (Table S1 in Supplementary Data). Reads were filtered based on quality using FASTX-Toolkit then PANDAseq [40] was used to assemble the read 1 and read 2 using the RDP algorithm (Ramer–Douglas–Peucker algorithm). The resulting sequences were annotated at the genus level by RDP Classifier [41] using the RDP 16srrna database and an assignment confidence cutoff of 0.6. The PCo (Principal coordinates) analysis (components 1 and 2 in (a), components 2 and 3 in (b)) of the Bray–Curtis dissimilarity matrix is based on the relative abundances of the different bacterial genera.

Figure 3.
Overview of the role of different factors in controlling microbial communities of the planetary boundary layer.
Figure 3.
Overview of the role of different factors in controlling microbial communities of the planetary boundary layer.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tignat-Perrier, R.; Dommergue, A.; Vogel, T.M.; Larose, C. Microbial Ecology of the Planetary Boundary Layer. Atmosphere 2020, 11, 1296. https://doi.org/10.3390/atmos11121296
AMA Style
Tignat-Perrier R, Dommergue A, Vogel TM, Larose C. Microbial Ecology of the Planetary Boundary Layer. Atmosphere. 2020; 11(12):1296. https://doi.org/10.3390/atmos11121296
Chicago/Turabian StyleTignat-Perrier, Romie, Aurélien Dommergue, Timothy M. Vogel, and Catherine Larose. 2020. "Microbial Ecology of the Planetary Boundary Layer" Atmosphere 11, no. 12: 1296. https://doi.org/10.3390/atmos11121296
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.