An Integrated Proteomic and Transcriptomic Analysis Reveals the Venom Complexity of the Bullet Ant Paraponera clavata

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. P. clavata Venom Gland Transcriptome Profile
2.1.1. Illumina Sequencing
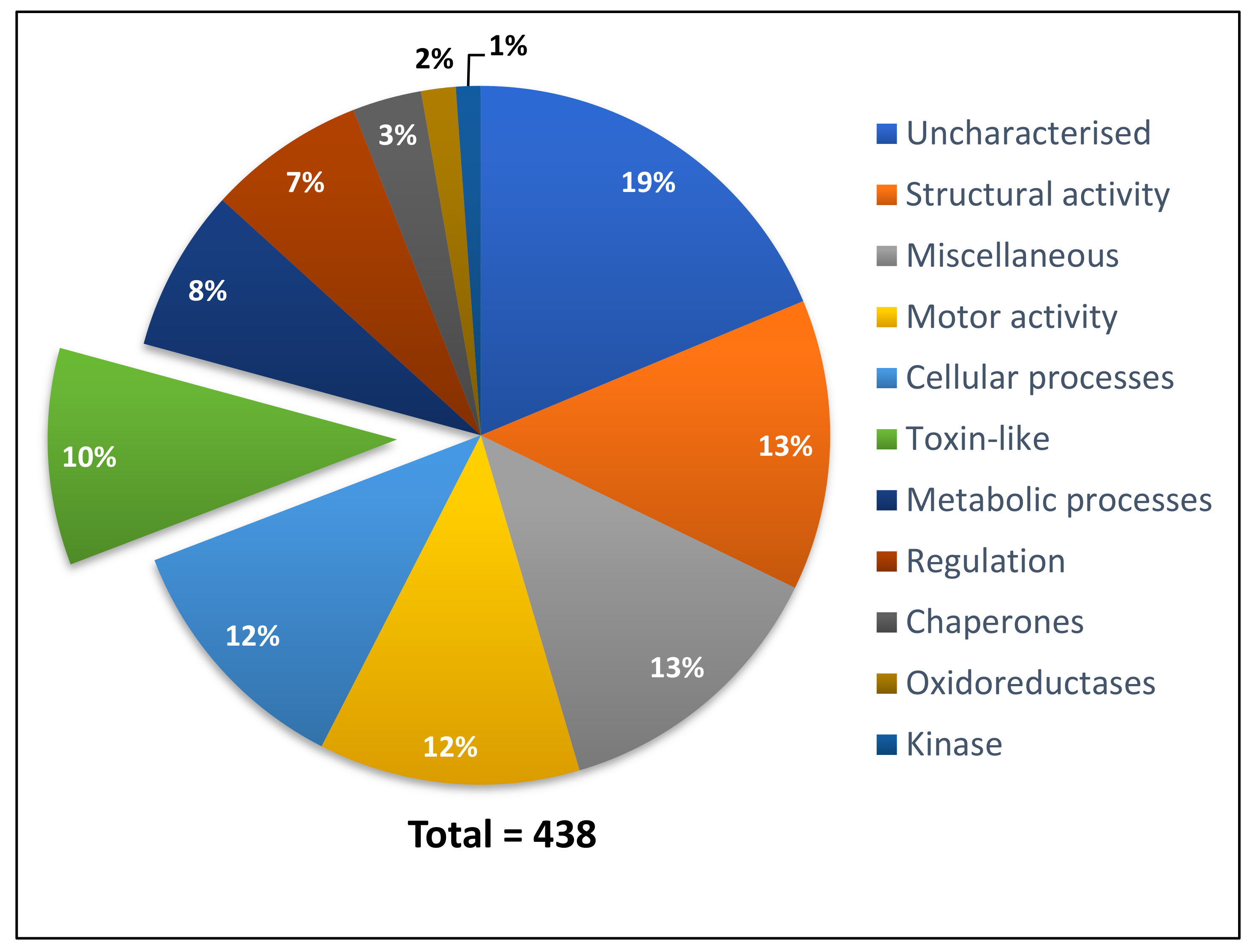
2.1.2. P. clavata Transcriptome Annotation
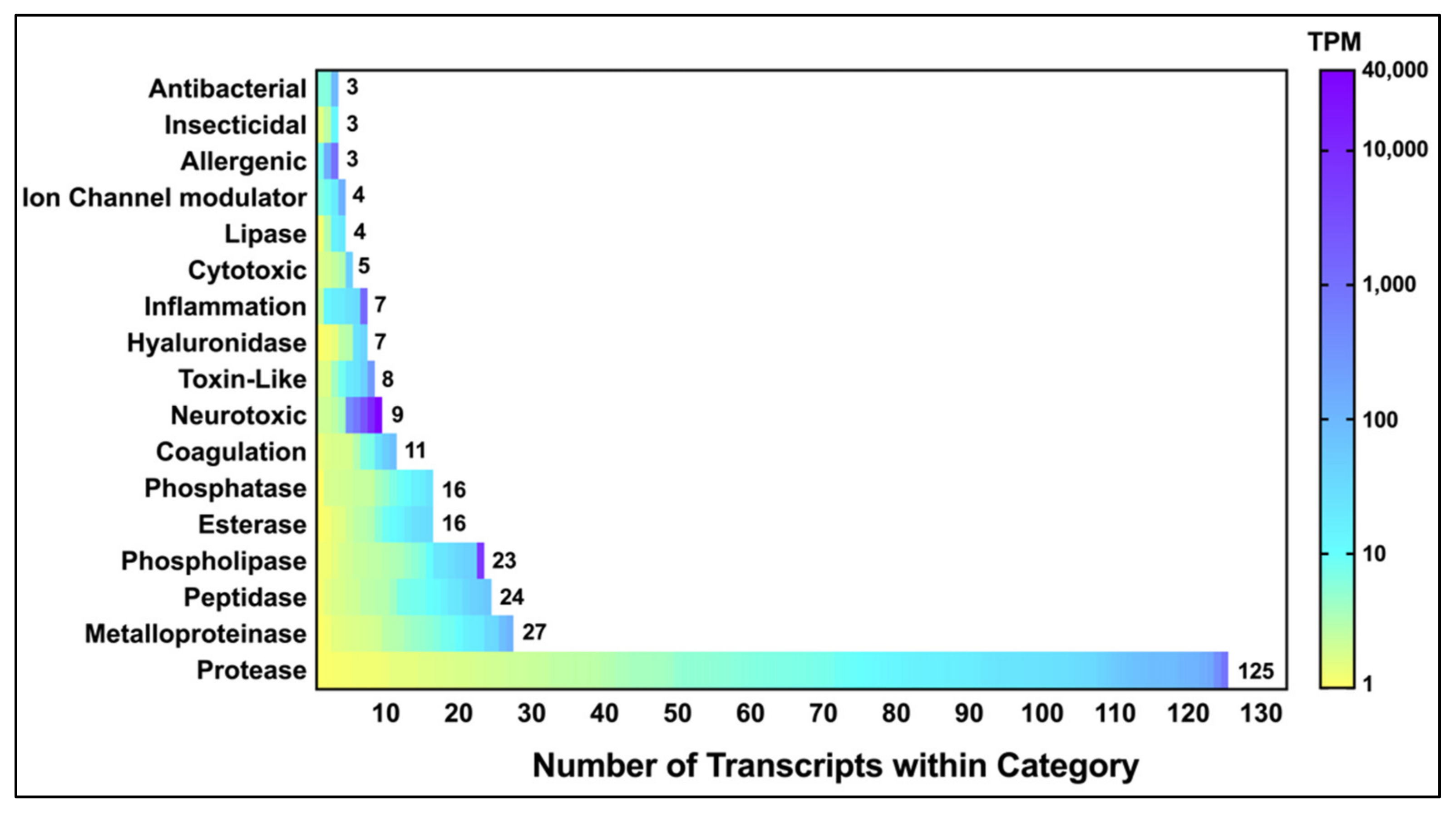
2.1.3. Identification and Classification of P. clavata Toxins
2.2. P. clavata Venom Proteome
2.3. P. clavata Toxins
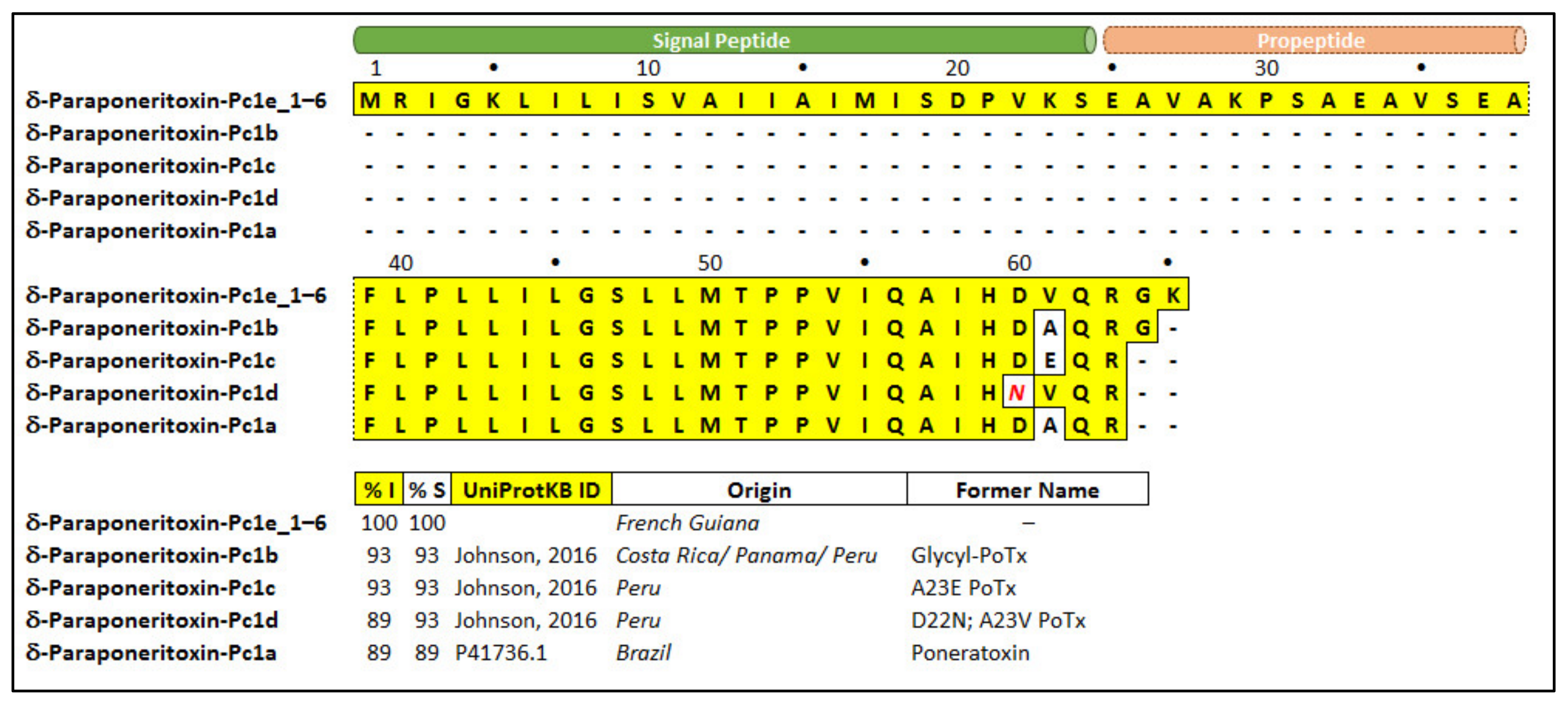
2.3.1. Poneratoxin
2.3.2. Inhibitor Cysteine Knot Peptide
2.3.3. Phospholipase A2 (PLA2)
2.3.4. Hyaluronidase
2.3.5. Icarapin
2.3.6. Arginine Kinase
2.3.7. Serine Proteases
2.4. Novel Toxin-Like Proteins
3. Discussion
3.1. P. clavata Transcriptome
3.1.1. Neurotoxins
3.1.2. Proteases
3.1.3. Phospholipases
3.1.4. Hyaluronidases
3.1.5. Allergenic Proteins
3.2. Protein and Peptide Transcripts Not Detected in the Venom Proteome
3.2.1. Toxin-Like Proteins
3.2.2. Insecticidal Proteins
3.2.3. Other Predicted P. clavata Toxin-Like Peptides
3.3. Combined Proteomic/ Transcriptomic Approach
4. Conclusions
5. Materials and Methods
5.1. Venom Gland Transcriptomics
5.2. Quality Control and de novo Transcriptome Assembly
5.3. Venom Gland Transcriptome Analysis
5.3.1. BLASTx
5.3.2. TransDecoder
5.3.3. Signal Peptide Prediction
5.3.4. Prediction of Toxin-Like Peptides
5.3.5. Toxin Identification
5.3.6. Nomenclature
5.3.7. Identification of Potential Pharmacological Targets
5.3.8. Transcriptome Data Submission
5.4. Venom Collection for Proteomics and Neuron Cell Assay
5.5. Protein Identification
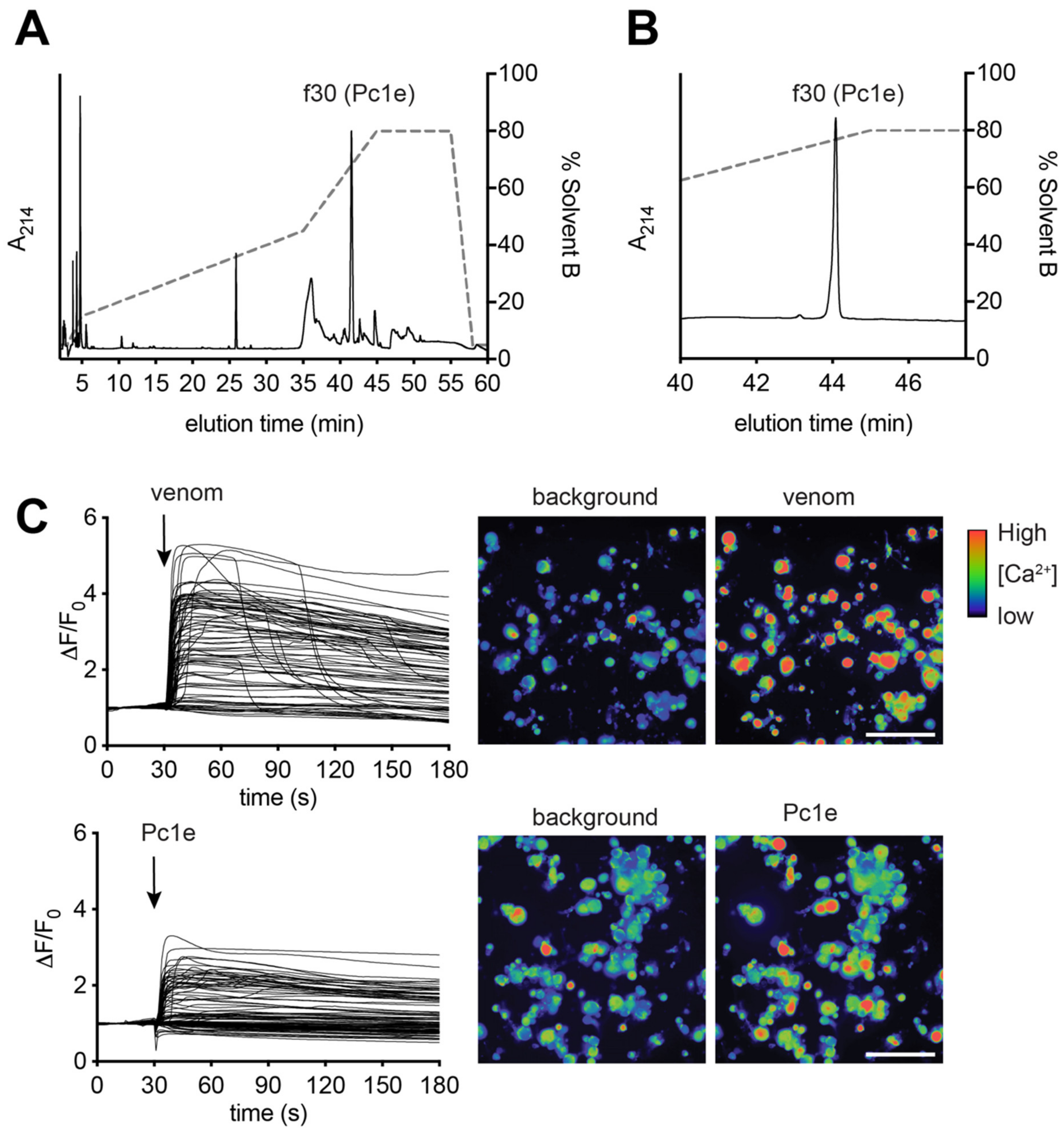
5.6. Purification of δ-paraponeritoxin-Pc1e by RP-HPLC
5.7. LC-MS/MS
5.8. Calcium Imaging of Sensory Neurons
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The diversity of venom: The importance of behavior and venom system morphology in understanding its ecology and evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.D.; Undheim, E.A.B.; Ueberheide, B.; King, G.F. Venom peptides as therapeutics: Advances, challenges and the future of venom-peptide discovery. Expert Rev. Proteom. 2017, 14, 931–939. [Google Scholar] [CrossRef]
- Smith, J.J.; Undheim, E.A.B. True lies: Using proteomics to assess the accuracy of transcriptome-based venomics in centipedes uncovers false positives and reveals startling intraspecific variation in Scolopendra subspinipes. Toxins 2018, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Piek, T.; Duval, A.; Hue, B.; Karst, H.; Lapied, B.; Mantel, P.; Nakajima, T.; Pelhate, M.; Schmidt, J.O. Poneratoxin, a novel peptide neurotoxin from the venom of the ant, Paraponera clavata. Comp. Biochem. Physiol. Part C Comp. Pharm. 1991, 99, 487–495. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mantel, P.; Nakajima, T.; Schmidt, J.O. Pharmacological characterization and chemical fractionation of the venom of the ponerine ant, Paraponera clavata (F.). Comp. Biochem. Physiol. Part C Comp. Pharm. 1991, 99, 481–486. [Google Scholar] [CrossRef]
- Duval, A.; Malécot, C.O.; Pelhate, M.; Piek, T. Poneratoxin, a new toxin from an ant venom, reveals an interconversion between two gating modes of the Na channels in frog skeletal muscle fibres. Pflügers Arch. 1992, 420, 239–247. [Google Scholar] [CrossRef]
- Hendrich, A.B.; Mozrzymas, J.W.; Konopińska, D.; Scuka, M. The effect of poneratoxin on neuromuscular transmission in the rat diaphragm. Cell. Mol. Biol. Lett. 2001, 7, 195–202. [Google Scholar]
- Szolajska, E.; Poznanski, J.; Ferber, M.L.; Michalik, J.; Gout, E.; Fender, P.; Bailly, I.; Dublet, B.; Chroboczek, J. Poneratoxin, a neurotoxin from ant venom. Structure and expression in insect cells and construction of a bio-insecticide. Eur. J. Biochem. 2004, 271, 2127–2136. [Google Scholar] [CrossRef]
- Rykaczewska-Czerwinska, M.; Radosz, A.; Konopinska, D.; Wrobel, M.; Plech, A. Antinociceptive effect of poneratoxin [PoTX] in rats. Pestycydy 2008, 1, 135–141. [Google Scholar]
- Johnson, S.R.; Rikli, H.G.; Schmidt, J.O.; Evans, M.S. A reexamination of poneratoxin from the venom of the bullet ant Paraponera clavata. Peptides 2017, 98, 51–62. [Google Scholar] [CrossRef]
- Aili, S.R.; Touchard, A.; Petitclerc, F.; Dejean, A.; Orivel, J.; Padula, M.P.; Escoubas, P.; Nicholson, G.M. Combined peptidomic and proteomic analysis of electrically stimulated and manually dissected venom from the south American bullet ant Paraponera clavata. J. Proteome Res. 2017, 16, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Aili, S.R.; Touchard, A.; Koh, J.M.S.; Dejean, A.; Orivel, J.; Padula, M.P.; Escoubas, P.; Nicholson, G.M. Comparisons of protein and peptide complexity in poneroid and formicoid ant venoms. J. Proteome Res. 2016, 15, 3039–3054. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from RNA-seq using the trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Pineda, S.S.; Chaumeil, P.-A.; Kunert, A.; Kaas, Q.; Thang, M.W.; Li, L.; Nuhn, M.; Herzig, V.; Saez, N.J.; Cristofori-Armstrong, B. ArachnoServer 3.0: An online resource for automated discovery, analysis and annotation of spider toxins. Bioinformatics 2017, 34, 1074–1076. [Google Scholar] [CrossRef] [Green Version]
- Graudins, A.; Little, M.J.; Pineda, S.S.; Hains, P.G.; King, G.F.; Broady, K.W.; Nicholson, G.M. Cloning and activity of a novel alpha-latrotoxin from red-back spider venom. Biochem. Pharm. 2012, 83, 170–183. [Google Scholar] [CrossRef] [Green Version]
- De Junqueira-de-Azevedo, L.M.I.; Pertinhez, T.; Spisni, A.; Carreño, F.R.; Farah, C.S.; Ho, P.L. Cloning and expression of calglandulin, a new EF-hand protein from the venom glands of Bothrops insularis snake in E. coli. Biochim. Biophys. Acta BBA Proteins Proteom. 2003, 1648, 90–98. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Piek, T. Neurotoxins from venoms of the hymenoptera-twenty-five years of research in Amsterdam. Comp. Biochem. Physiol. Part C Comp. Pharm. 1990, 96, 223–233. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- McCleskey, E.; Fox, A.; Feldman, D.; Cruz, L.; Olivera, B.; Tsien, R.; Yoshikami, D. Omega-conotoxin: Direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Proc. Natl. Acad. Sci. USA 1987, 84, 4327–4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.M.; Lomonte, B. Phospholipases A2: Unveiling the secrets of a functionally versatile group of snake venom toxins. Toxicon 2013, 62, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.L.; Otwinowski, Z.; Gelb, M.H.; Sigler, P.B. Crystal structure of bee-venom phospholipase A2 in a complex with a transition-state analogue. Science 1990, 250, 1563–1566. [Google Scholar] [CrossRef] [Green Version]
- Six, D.A.; Dennis, E.A. The expanding superfamily of phospholipase A2 enzymes: Classification and characterization. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2000, 1488, 1–19. [Google Scholar] [CrossRef]
- Valentin, E.; Ghomashchi, F.; Gelb, M.H.; Lazdunski, M.; Lambeau, G. Novel human secreted phospholipase A2 with homology to the group III bee venom enzyme. J. Biol. Chem. 2000, 275, 7492–7496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuchler, K.; Gmachl, M.; Sippl, M.J.; Kreil, G. Analysis of the cDNA for phospholipase A2 from honeybee venom glands. Eur. J. Biochem. 1989, 184, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, W.; Verdenaud, M.; Klopp, C.; Ducancel, F.; Noirot, C.; Vétillard, A. De novo sequencing and transcriptome analysis for Tetramorium bicarinatum: A comprehensive venom gland transcriptome analysis from an ant species. BMC Genom. 2014, 15, 987. [Google Scholar] [CrossRef] [Green Version]
- White, S.P.; Scott, D.L.; Otwinowski, Z.; Gelb, M.H.; Sigler, P.B. Crystal structure of cobra-venom phospholipase A2 in a complex with a transition-state analogue. Science 1990, 250, 1560–1563. [Google Scholar] [CrossRef]
- Kini, R.M. Structure-function relationships and mechanism of anticoagulant phospholipase A 2 enzymes from snake venoms. Toxicon 2005, 45, 1147–1161. [Google Scholar] [CrossRef]
- Chioato, L.; Ward, R.J. Mapping structural determinants of biological activities in snake venom phospholipases A2 by sequence analysis and site directed mutagenesis. Toxicon 2003, 42, 869–883. [Google Scholar] [CrossRef]
- Marković-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Müller, U.; Schirmer, T. Crystal structure of hyaluronidase, a major allergen of bee venom. Structure 2000, 8, 1025–1035. [Google Scholar] [CrossRef]
- Peiren, N.; de Graaf, D.C.; Brunain, M.; Bridts, C.H.; Ebo, D.G.; Stevens, W.J.; Jacobs, F.J. Molecular cloning and expression of icarapin, a novel IgE-binding bee venom protein. FEBS Lett. 2006, 580, 4895–4899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, J.H.; Lee, S.H. Identification and characterization of venom proteins of two solitary wasps, Eumenes pomiformis and Orancistrocerus drewseni. Toxicon 2010, 56, 554–562. [Google Scholar] [CrossRef]
- Freeman, T.M.; Tracy, J.M. Stinging Insect Allergy: A Clinician’s Guide; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Schmidt, J.O. The Sting of the Wild; Johns Hopkins University Press: Baltimore, MD, USA, 2016. [Google Scholar]
- Torres, A.F.C.; Huang, C.; Chong, C.-M.; Leung, S.W.; Prieto-da-Silva, A.R.B.; Havt, A.; Quinet, Y.P.; Martins, A.M.C.; Lee, S.M.Y.; Rádis-Baptista, G. Transcriptome analysis in venom gland of the predatory giant ant Dinoponera quadriceps: Insights into the polypeptide toxin arsenal of Hymenopterans. PLoS ONE 2014, 9, e87556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponce, D.; Brinkman, D.L.; Potriquet, J.; Mulvenna, J. Tentacle transcriptome and venom proteome of the pacific sea nettle, Chrysaora fuscescens (Cnidaria: Scyphozoa). Toxins 2016, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsik, C.G.; Tayal, A.; Diesh, C.M.; Unni, D.R.; Emery, M.L.; Nguyen, H.N.; Hagen, D.E. Hymenoptera Genome Database: Integrating genome annotations in HymenopteraMine. Nucleic Acids Res. 2016, 44, D793–D800. [Google Scholar] [CrossRef] [Green Version]
- Aili, S.R.; Touchard, A.; Escoubas, P.; Padula, M.P.; Orivel, J.; Dejean, A.; Nicholson, G.M. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef]
- Moreau, C.S.; Bell, C.D.; Vila, R.; Archibald, S.B.; Pierce, N.E. Phylogeny of the ants: Diversification in the age of angiosperms. Science 2006, 312, 101–104. [Google Scholar] [CrossRef]
- Robinson, S.D.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.R.; Payne, R.J.; Vetter, I.; King, G.F.; Undheim, E.A. A comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4, 4640. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, S.; Malyavka, A.; McCutchen, B.; Lu, A.; Schepers, E.; Herrmann, R.; Grishin, E. A novel strategy for the identification of toxin-like structures in spider venom. Proteins Struct. Funct. Bioinform. 2005, 59, 131–140. [Google Scholar] [CrossRef]
- Wong, E.S.; Hardy, M.C.; Wood, D.; Bailey, T.; King, G.F. SVM-based prediction of propeptide cleavage sites in spider toxins identifies toxin innovation in an Australian tarantula. PLoS ONE 2013, 8, e66279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouls, L.; Subra, G.; Aubagnac, J.L.; Martinez, J.; Enjalbal, C. Tandem mass spectrometry of amidated peptides. J. Mass Spectrom. 2006, 41, 1470–1483. [Google Scholar] [CrossRef] [PubMed]
- Labella, C.; Kanawati, B.; Vogel, H.; Schmitt-Kopplin, P.; Laurino, S.; Bianco, G.; Falabella, P. Identification of two arginine kinase forms of endoparasitoid Leptomastix dactylopii venom by bottom up-sequence tag approach. J. Mass Spectrom. 2015, 50, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Arimoto, H.; Kinumi, T.; Oba, Y.; Uemura, D. Identification of proteins from venom of the paralytic spider wasp, Cyphononyx dorsalis. Insect Biochem. Mol. Biol. 2007, 37, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Kim, B.Y.; Sohn, M.R.; Roh, J.Y.; Je, Y.H.; Kim, N.J.; Kim, I.; Woo, S.D. Dual function of a bee venom serine protease: Prophenoloxidase-activating factor in arthropods and fibrin(ogen)olytic enzyme in mammals. PLoS ONE 2010, 5, e10393. [Google Scholar] [CrossRef]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochim. Biophys. Acta BBA Protein Struct. Mol. Enzymol. 2000, 1477, 146–156. [Google Scholar] [CrossRef]
- Jiang, H.; Kanost, M.R. The clip-domain family of serine proteinases in arthropods. Insect Biochem. Mol. Biol. 2000, 30, 95–105. [Google Scholar] [CrossRef]
- Winningham, K.M.; Fitch, C.D.; Schmidt, M.; Hoffman, D.R. Hymenoptera venom protease allergens. J. Allergy Clin. Immunol. 2004, 114, 928–933. [Google Scholar] [CrossRef]
- Laurino, S.; Grossi, G.; Pucci, P.; Flagiello, A.; Bufo, S.A.; Bianco, G.; Salvia, R.; Vinson, S.B.; Vogel, H.; Falabella, P. Identification of major Toxoneuron nigriceps venom proteins using an integrated transcriptomic/proteomic approach. Insect Biochem. Mol. Biol. 2016, 76, 49–61. [Google Scholar] [CrossRef]
- Asgari, S.; Zhang, G.; Zareie, R.; Schmidt, O. A serine proteinase homolog venom protein from an endoparasitoid wasp inhibits melanization of the host hemolymph. Insect Biochem. Mol. Biol. 2003, 33, 1017–1024. [Google Scholar] [CrossRef]
- Ross, J.; Jiang, H.; Kanost, M.R.; Wang, Y. Serine proteases and their homologs in the Drosophila melanogaster genome: An initial analysis of sequence conservation and phylogenetic relationships. Gene 2003, 304, 117–131. [Google Scholar] [CrossRef]
- Piao, S.; Song, Y.L.; Kim, J.H.; Park, S.Y.; Park, J.W.; Lee, B.L.; Oh, B.H.; Ha, N.C. Crystal structure of a clip-domain serine protease and functional roles of the clip domains. EMBO J. 2005, 24, 4404–4414. [Google Scholar] [CrossRef] [PubMed]
- Kazuma, K.; Masuko, K.; Konno, K.; Inagaki, H. Combined venom gland transcriptomic and venom peptidomic analysis of the predatory ant Odontomachus monticola. Toxins 2017, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Luna-Ramírez, K.; Quintero-Hernández, V.; Juárez-González, V.R.; Possani, L.D. Whole transcriptome of the venom gland from Urodacus yaschenkoi scorpion. PLoS ONE 2015, 10, e0127883. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase a 2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef]
- Jacomini, D.L.J.; Pereira, F.D.C.; dos Santos Pinto, J.R.A.; dos Santos, L.D.; da Silva Neto, A.J.; Giratto, D.T.; Palma, M.S.; de Lima Zollner, R.; Braga, M.R.B. Hyaluronidase from the venom of the social wasp Polybia paulista (Hymenoptera, Vespidae): Cloning, structural modeling, purification, and immunological analysis. Toxicon 2013, 64, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Focke, M.; Léonard, R.; Jarisch, R.; Altmann, F.; Hemmer, W. Reassessing the role of hyaluronidase in yellow jacket venom allergy. J. Allergy Clin. Immunol. 2010, 125, 184–190. [Google Scholar] [CrossRef]
- Hoffman, D.R. Hymenoptera venom allergens. Clin. Rev. Allergy Immunol. 2006, 30, 109–128. [Google Scholar] [CrossRef]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [Green Version]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Zobel-Thropp, P.A.; Kerins, A.E.; Binford, G.J. Sphingomyelinase D in sicariid spider venom is a potent insecticidal toxin. Toxicon 2012, 60, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Binford, G.J.; Cordes, M.H.; Wells, M.A. Sphingomyelinase D from venoms of Loxosceles spiders: Evolutionary insights from cDNA sequences and gene structure. Toxicon 2005, 45, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Ueberheide, B.M.; Fenyö, D.; Alewood, P.F.; Chait, B.T. Rapid sensitive analysis of cysteine rich peptide venom components. Proc. Natl. Acad. Sci. USA 2009, 106, 6910–6915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, G.; Cohen, M. The expression and phylogenetics of the inhibitor cysteine knot peptide OCLP1 in the honey bee Apis mellifera. J. Insect Physiol. 2014, 65, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Arias, H.R.; Blanton, M.P. α-Conotoxins. Int. J. Biochem. Cell Biol. 2000, 32, 1017–1028. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.H.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [Green Version]
- Lader, E.S. Methods and Reagents for Preserving Rna in Cell and Tissue Samples; Patent and Trademark Office: Washington, DC, USA, 2012. [Google Scholar]
- Andrews, S. FastQC: A quality control tool for high throughput sequence data; Babraham Bioinformatics Group: Cambridge, UK, USA, 2011. [Google Scholar]
- Lohse, M.; Bolger, A.M.; Nagel, A.; Fernie, A.R.; Lunn, J.E.; Stitt, M.; Usadel, B. R obi NA: A user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 2012, 40, W622–W627. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Consortium, U. The universal protein resource (UniProt). Nucleic Acids Res. 2008, 36, D190–D195. [Google Scholar] [CrossRef]
- Huntley, R.P.; Sawford, T.; Mutowo-Meullenet, P.; Shypitsyna, A.; Bonilla, C.; Martin, M.J.; O’Donovan, C. The GOA database: Gene ontology annotation updates for 2015. Nucleic Acids Res. 2015, 43, D1057–D1063. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.a.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Touchard, A.; Aili, S.; Fox, E.; Escoubas, P.; Orivel, J.; Nicholson, G.; Dejean, A. The biochemical toxin arsenal from ant venoms. Toxins 2016, 8, 30. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| After QC | Counts |
|---|---|
| Total number of reads (paired end) | 17,026,962 |
| Assembly summary | |
| Total number of contigs | 54,242 |
| GC content | 37.3% |
| Raw reads mapped to contigs | 99.3% |
| Annotation | |
| Total BLASTx hits | 37,140 |
| Assembled contigs with BLASTx hits | 32, 003 |
| Contigs with gene ontology annotation | 6732 |
| At least one ORF | 31,586 |
| * Toxins | 295 |
| Proportion of contigs that are toxins | 4.4% |
| Toxin Name | BLASTx/ Tox|Note Protein Name | NCBInr Accession | Species | Trinity ID | TPM | Toxin Category |
|---|---|---|---|---|---|---|
| δ-Paraponeritoxin-Pc1e_1 | Poneratoxin | P41736.1 | Paraponera clavata | DN10254_c0_g26_i11 | 39,657 | Neurotoxic |
| δ-Paraponeritoxin-Pc1e_2 | DN9710_c0_g11_i2 | 9279 | ||||
| δ-Paraponeritoxin-Pc1e_3 | DN10254_c0_g26_i12 | 2414 | ||||
| δ-Paraponeritoxin-Pc1e_4 | DN10254_c0_g26_i5 | 797 | ||||
| Phospholipase-A2-1a_1-P-clavata | PREDICTED: phospholipase A2 | XP_011262712 | Camponotus floridanus | DN9894_c0_g8_i1 | 6328 | Phospholipase |
| Translationally-controlled-tumor-protein-1a-P-clavata | PREDICTED: translationally controlled tumour protein homolog | XP_014489127 | Dinoponera quadriceps | DN9555_c0_g10_i3 | 1229 | Inflammation |
| Icarapin-1a-P-clavata | PREDICTED: icarapin-like | XP_011863857 | Vollenhovia emeryi | DN9969_c1_g1_i4 | 969 | Allergenic |
| Serine-protease-1a-P-clavata | Serine proteinase stubble | EZA53191.1 | Ooceraea biroi | DN10218_c2_g11_i1 | 835 | Protease |
| Serine-protease-2a-P-clavata | DN10218_c2_g7_i1 | 364 | ||||
| Arginine-kinase-1a_1-P-clavata | PREDICTED: arginine kinase isoform X1 | XP_014467324 | Dinoponera quadriceps | DN9942_c0_g1_i6 | 382 | Neurotoxic |
| U1-Paraponeritoxin-Pc1a | PREDICTED: omega-conotoxin-like protein 1 | XP_011257508 | Camponotus floridanus | DN5808_c0_g1_i1 | 253 | Unknown |
| Serine-protease-3a-P-clavata | PREDICTED: serine protease easter-like isoform X2 | XP_014488937 | Dinoponera quadriceps | DN10218_c1_g1_i2 | 161 | Protease |
| Serine-protease-4a-P-clavata | Serine protease snake | KMR05322.1 | Lasius niger | DN11547_c0_g1_i1 | 138 | Protease |
| Venom-allergen3-1a-P-clavata | PREDICTED: venom allergen 3-like | XP_012534360 | Monomorium pharaonis | DN10202_c0_g4_i2 | 127 | Allergenic |
| Cysteine-rich protein-1a-P-clavata | Cysteine-rich protein 1 | KYN02327.1 | Cyphomyrmex costatus | DN7413_c0_g4_i1 | 123 | Ion channel modulator |
| Serine-protease-1b-P-clavata | PREDICTED: serine protease 42-like isoform X3 | XP_014488938 | Dinoponera quadriceps | DN10218_c2_g8_i1 | 113 | Protease |
| Serine-protease-5a-P-clavata | DN10218_c2_g6_i7 | 103 | ||||
| Disintegrin-and-metalloproteinase-domain-containing-protein-1a-P-clavata | Disintegrin and metalloproteinase domain-containing protein | XP_012242558 | Bombus impatiens | DN21256_c0_g1_i1 | 113 | Metalloproteinase |
| Disintegrin-and-metalloproteinase-domain-containing-protein-2a-P-clavata | EZA52689.1 | Ooceraea biroi | DN20154_c0_g1_i1 | 75 | ||
| Lysosomal-aspartic-protease-1a-P-clavata | PREDICTED: lysosomal aspartic protease | XP_014468660 | Dinoponera quadriceps | DN7128_c0_g1_i1 | 111 | Protease |
| Defensin-2-like-1a-P-clavata | PREDICTED: defensin-2-like | XP_012537462 | Monomorium pharaonis | DN5917_c0_g1_i1 | 89 | Antibacterial |
| Serine-protease-inhibitor-1a-P-clavata | PREDICTED: serine protease inhibitor 3/4-like isoform X2 | XP_012539061 | Monomorium pharaonis | DN9965_c0_g10_i1 | 86 | Protease |
| Serine-protease-6a-P-clavata | PREDICTED: venom serine protease-like | XP_014477136 | Dinoponera quadriceps | DN9366_c0_g3_i1 | 85 | Protease |
| Serine-protease-7a-P-clavata | PREDICTED: serine protease 52-like | XP_011641802 | Pogonomyrmex barbatus | DN10218_c2_g11_i2 | 84 | Protease |
| Serine-protease-8a-P-clavata | PREDICTED: serine protease snake-like | XP_011142823 | Harpegnathos saltator | DN9987_c0_g2_i9 | 83 | Protease |
| Cathepsin-L-1a-P-clavata | Cathepsin L | EFN75465.1 | Harpegnathos saltator | DN9254_c0_g3_i1 | 77 | Protease |
| Toxin Name | BLASTx Hit | Species | Trinity ID | Unique Peptide | % Cover | −10lgP | TPM | SigP (y/n) |
|---|---|---|---|---|---|---|---|---|
| Hyalyronidase-1a_1-P-clavata | PREDICTED: hyaluronidase-like | D. quadriceps | DN10062_c0_g13_i9 | 70 | 65 | 333 | 38 | Y |
| Hyalyronidase-1a_2-P-clavata | DN10062_c0_g13_i12 | 70 | 65 | 333 | 23 | Y | ||
| Hyalyronidase-1a_3-P-clavata | DN10062_c0_g13_i7 | 70 | 65 | 333 | 3 | Y | ||
| Hyalyronidase-1a_4-P-clavata | DN10062_c0_g13_i16 | 70 | 65 | 333 | 3 | Y | ||
| Hyalyronidase-1a_5-P-clavata | DN10062_c0_g13_i15 | 70 | 65 | 333 | 1 | Y | ||
| Hyalyronidase-1a_6-P-clavata | DN10062_c0_g13_i3 | 70 | 65 | 333 | 1 | Y | ||
| Hyalyronidase-1a_7-P-clavata | DN10062_c0_g13_i10 | 70 | 65 | 333 | 1 | Y | ||
| Hyalyronidase-1a_8-P-clavata | DN10062_c0_g13_i5 | 70 | 65 | 333 | 1 | Y | ||
| Venom-dipeptidyl-peptidase-1a_1-P-clavata | PREDICTED: venom dipeptidyl peptidase 4 | L. humile | DN8858_c0_g1_i1 | 26 | 29 | 281 | 60 | N |
| Venom-dipeptidyl-peptidase-1a_2-P-clavata | PREDICTED: venom dipeptidyl peptidase 4 | DN8858_c0_g1_i2 | 26 | 29 | 281 | 21 | N | |
| δ-Paraponeritoxin-Pc1e_2 | Poneratoxin | P. clavata | DN9710_c0_g11_i2 | 20 | 45 | 260 | 9279 | Y |
| δ-Paraponeritoxin-Pc1e_5 | DN10254_c0_g26_i8 | 20 | 45 | 260 | 2 | Y | ||
| δ-Paraponeritoxin-Pc1e_6 | DN10254_c0_g26_i3 | 20 | 45 | 260 | 0 | Y | ||
| Phospholipase-A2-1a_1-P-clavata | PREDICTED: phospholipase A2 | C. floridanus | DN9894_c0_g8_i1 | 24 | 48 | 235 | 6328 | Y |
| Phospholipase-A2-1a_2-P-clavata | DN9894_c0_g2_i2 | 24 | 48 | 235 | 7 | Y | ||
| Venom-allergen3-1a-P-clavata | PREDICTED: venom allergen 3-like | M. pharaonis | DN10202_c0_g4_i2 | 14 | 44 | 198 | 127 | Y |
| Venom-allergen3-1b-P-clavata | DN10202_c0_g4_i1 | 14 | 53 | 198 | 1 | Y | ||
| Carboxypeptidase-Q-1a_1-P-clavata | PREDICTED: carboxypeptidase Q | C. biroi | DN8288_c0_g1_i2 | 12 | 22 | 180 | 3 | N |
| Carboxypeptidase-Q-1a_2-P-clavata | DN8288_c0_g1_i1 | 12 | 23 | 180 | 2 | N | ||
| Arginine-kinase-1a_1-P-clavata | PREDICTED: arginine kinase isoform X1 | D. quadriceps | DN9942_c0_g1_i6 | 10 | 32 | 169 | 382 | Y |
| Arginine-kinase-1a_2-P-clavata | DN9942_c0_g1_i1 | 10 | 32 | 169 | 4 | N | ||
| Pancreatic-lipase-1a-P-clavata | Pancreatic lipase-related protein 2 | C. floridanus | DN10238_c1_g31_i1 | 8 | 23 | 169 | 15 | Y |
| Pancreatic-lipase-1b-P-clavata | DN10238_c1_g10_i1 | 8 | 25 | 169 | 3 | Y | ||
| Serine-protease-6a-P-clavata | PREDICTED: venom serine protease-like | D. quadriceps | DN9366_c0_g3_i1 | 6 | 10 | 135 | 85 | Y |
| Matrix-metalloproteinase-14-like-1a-P-clavata | Matrix metalloproteinase-14 | C. biroi | DN18586_c0_g1_i1 | 3 | 23 | 104 | 5 | N |
| Serine-protease-inhibitor-1a-P-clavata | PREDICTED: serine protease inhibitor 3/4-like isoform X2 | M. pharaonis | DN9965_c0_g10_i1 | 4 | 11 | 97 | 86 | Y |
| Alaserpin-X5-like-1a-P-clavata | PREDICTED: alaserpin-like isoform X5 | P. barbatus | DN9965_c0_g10_i2 | 4 | 12 | 97 | 18 | Y |
| Pancreatic-triacylglycerol-lipase-X1-like-1a-P-clavata | PREDICTED: pancreatic triacylglycerol lipase-like isoform X1 | D. quadriceps | DN8997_c0_g2_i2 | 3 | 6 | 93 | 5 | Y |
| Pancreatic-triacylglycerol-lipase-X1-like-2a-P-clavata | DN8997_c0_g2_i3 | 1 | 3 | 37 | 2 | Y | ||
| Pancreatic-triacylglycerol-lipase-X1-like-3a-P-clavata | DN8997_c0_g2_i1 | 1 | 8 | 37 | 1 | Y | ||
| Venom-acid-phosphatase-1a-P-clavata | PREDICTED: venom acid phosphatase Acph-1 | C. biroi | DN5022_c0_g1_i1 | 3 | 7 | 63 | 5 | N |
| Iron-zinc-purple-acid phosphatase-like-1a_1-P-clavata | Iron zinc purple acid phosphatase-like protein | L. niger | DN8672_c0_g1_i2 | 2 | 4 | 55 | 17 | N |
| Iron-zinc-purple-acid phosphatase-like-1a_2-P-clavata | DN8672_c0_g1_i1 | 2 | 4 | 55 | 9 | N | ||
| Alkaline-phosphatase-1a_1-P-clavata | PREDICTED: alkaline phosphatase 4-like | L. humile | DN9670_c0_g1_i1 | 1 | 2 | 51 | 24 | N |
| Alkaline-phosphatase-1a_2-P-clavata | Alkaline phosphatase 4 | H. saltator | DN9670_c0_g1_i2 | 1 | 2 | 51 | 2.3 | N |
| Neprilysin-11-isoform-X2-1a_1-P-clavata | PREDICTED: neprilysin-11 isoform X2 | D. quadriceps | DN10125_c0_g9_i1 | 2 | 4 | 43 | 4.5 | Y |
| Neprilysin-11--isoform-X2-1a_2-P-clavata | PREDICTED: neprilysin-11 isoform X2 | DN10125_c0_g18_i1 | 2 | 4 | 43 | 1.7 | Y | |
| Matrix-metalloproteinase-14-like-2a-P-clavata | PREDICTED: matrix metalloproteinase-14 isoform | D. quadriceps | DN21699_c0_g1_i1 | 2 | 6 | 41 | 1.9 | Y |
| Protein-5NUC-1a-P-clavata | PREDICTED: protein 5NUC-like | Wasmannia auropunctata | DN10097_c1_g11_i1 | 1 | 2 | 37 | 25.2 | N |
| Icarapin-1a-P-clavata | PREDICTED: icarapin-like | D. quadriceps | DN9969_c1_g1_i4 | 1 | 7 | 32 | 969 | N |
| Cytosolic-dipeptidase-1a-P-clavata | PREDICTED: cytosolic non-specific dipeptidase | D. quadriceps | DN9768_c0_g1_i2 | 1 | 2 | 30 | 1.8 | Y |
| Phenoloxidase-2-like-1a-P-clavata | PREDICTED: phenoloxidase 2 | S. invicta | DN15311_c0_g1_i1 | 1 | 4 | 24 | 3.2 | Y |
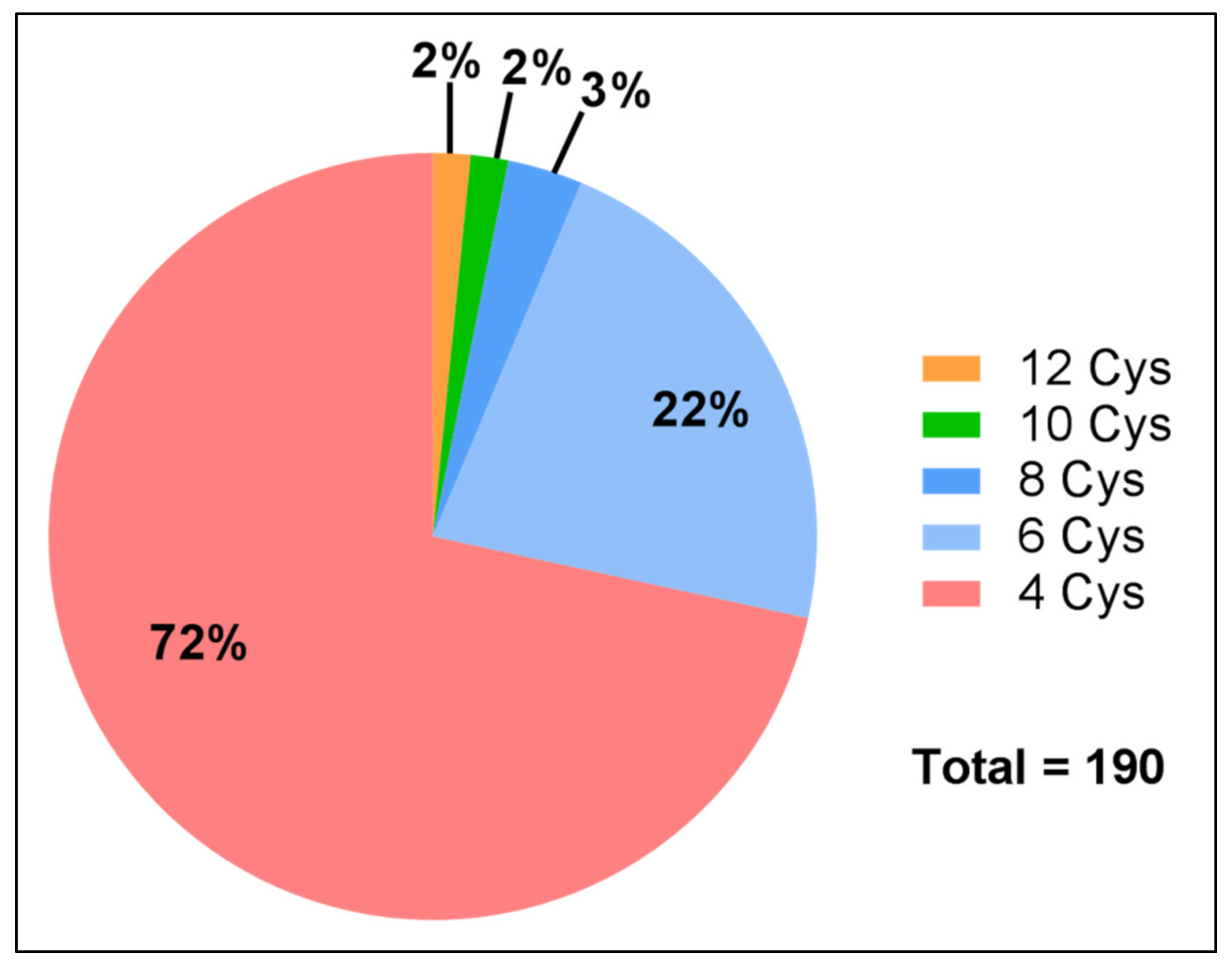
| Number of Cysteines | Scaffold | Mature Peptide Length | TPM |
|---|---|---|---|
| 4 | x8CCx1Cx19Cx5 | 37 | 772 |
| x1CCx34Cx1Cx1 | 41 | 18.4 | |
| x5CCx1Cx16Cx14 | 40 | 16.5 | |
| CCx23Cx7Cx6 | 40 | 13 | |
| x14CCx11Cx7Cx33 | 69 | 10.8 | |
| CCx1Cx27Cx12 | 44 | 7.1 | |
| x3CCx6Cx15Cx9 | 37 | 2.2 | |
| x6CCx7Cx11Cx8 | 36 | 0.76 | |
| x8CCx5Cx12Cx6 | 35 | 0.3 | |
| 6 | Cx6Cx5CCx4Cx6Cx29 | 56 | 43.8 |
| x18Cx6Cx3CC8Cx24Cx7 | 72 | 9.3 | |
| x7Cx8Cx21CCx5Cx22Cx20 | 89 | 5.9 | |
| Cx6Cx5CCx4Cx6Cx29 | 56 | 3.9 | |
| x8Cx6Cx5CCx4Cx6Cx25 | 60 | 3.4 | |
| x8Cx6Cx5CCx4Cx6Cx7 | 42 | 2.6 | |
| 8 | x14Cx9Cx3Cx5Cx5CCx3Cx4Cx26 | 77 | 1.76 |
| 10 | x10Cx5Cx4CCx11Cx9Cx28Cx1Cx5Cx6Cx1 | 90 | 12.67 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aili, S.R.; Touchard, A.; Hayward, R.; Robinson, S.D.; Pineda, S.S.; Lalagüe, H.; Mrinalini; Vetter, I.; Undheim, E.A.B.; Kini, R.M.; et al. An Integrated Proteomic and Transcriptomic Analysis Reveals the Venom Complexity of the Bullet Ant Paraponera clavata. Toxins 2020, 12, 324. https://doi.org/10.3390/toxins12050324
Aili SR, Touchard A, Hayward R, Robinson SD, Pineda SS, Lalagüe H, Mrinalini, Vetter I, Undheim EAB, Kini RM, et al. An Integrated Proteomic and Transcriptomic Analysis Reveals the Venom Complexity of the Bullet Ant Paraponera clavata. Toxins. 2020; 12(5):324. https://doi.org/10.3390/toxins12050324
Chicago/Turabian StyleAili, Samira R., Axel Touchard, Regan Hayward, Samuel D. Robinson, Sandy S. Pineda, Hadrien Lalagüe, Mrinalini, Irina Vetter, Eivind A. B. Undheim, R. Manjunatha Kini, and et al. 2020. "An Integrated Proteomic and Transcriptomic Analysis Reveals the Venom Complexity of the Bullet Ant Paraponera clavata" Toxins 12, no. 5: 324. https://doi.org/10.3390/toxins12050324